Potrebbero piacerti anche
- Riassunti Di Semeiotica MedicaDocumento48 pagineRiassunti Di Semeiotica Medicaantonio nenna100% (4)
- Autismo Intestino Alimentazione Sistema ImmunitarioDa EverandAutismo Intestino Alimentazione Sistema ImmunitarioNessuna valutazione finora
- BATTERIOLOGIA (Antonelli-Microbiologia Medica)Documento10 pagineBATTERIOLOGIA (Antonelli-Microbiologia Medica)Martina AntinozziNessuna valutazione finora
- Chirurgica 1° Sem - Prelievi Ed EmocoltureDocumento9 pagineChirurgica 1° Sem - Prelievi Ed Emocoltureantonio nennaNessuna valutazione finora
- La Sindrome del Colon Irritabile: Guida alla fisiopatologia, alimentazione ed integrazioneDa EverandLa Sindrome del Colon Irritabile: Guida alla fisiopatologia, alimentazione ed integrazioneNessuna valutazione finora
- Sindrome Hemolitico UremicoDocumento36 pagineSindrome Hemolitico Uremicojose vattuonoNessuna valutazione finora
- Miv Patogenesi Delle Infezioni BattericheDocumento9 pagineMiv Patogenesi Delle Infezioni Battericheantonio nennaNessuna valutazione finora
- PancreatitiDocumento64 paginePancreatitiapi-3733154100% (1)
- Miv Patogenesi Delle Infezioni BattericheDocumento7 pagineMiv Patogenesi Delle Infezioni Battericheantonio nennaNessuna valutazione finora
- 2 Obbiettivi Risposta Infiammatoria PDFDocumento21 pagine2 Obbiettivi Risposta Infiammatoria PDFAndrea Il MiodoNessuna valutazione finora
- Patologia GeneraleDocumento7 paginePatologia GeneraleCindy PhillipsNessuna valutazione finora
- Biologia Cellulare - 13 ApoptosiDocumento29 pagineBiologia Cellulare - 13 ApoptosiGuenhNessuna valutazione finora
- Lezione Piastrinopenie e Piastrinosi 2023Documento58 pagineLezione Piastrinopenie e Piastrinosi 2023rcalzd00Nessuna valutazione finora
- Biologia Applicata - ApoptosiDocumento30 pagineBiologia Applicata - Apoptosiapi-3826003100% (4)
- Lezione 5 - Morte CellulareDocumento107 pagineLezione 5 - Morte CellularelucaNessuna valutazione finora
- 05 Fegato 1Documento67 pagine05 Fegato 1Renzo LigurgoNessuna valutazione finora
- EcoliDocumento62 pagineEcoliapi-3863584Nessuna valutazione finora
- Domande Esame MicrobiologiaDocumento5 pagineDomande Esame MicrobiologiamichNessuna valutazione finora
- MorfopatoDocumento110 pagineMorfopatocrisssssssssNessuna valutazione finora
- 2021 FARMACIA Istologia 5 Sangue 12 MarzoDocumento52 pagine2021 FARMACIA Istologia 5 Sangue 12 MarzoChiara ArdauNessuna valutazione finora
- Capitolo 23 Controllo Delle Malattie InfettiveDocumento6 pagineCapitolo 23 Controllo Delle Malattie InfettiveClaudia Di finizioNessuna valutazione finora
- 12) Calcio e Altri Ti NVDocumento29 pagine12) Calcio e Altri Ti NVapi-26155364Nessuna valutazione finora
- Sadava Soluzioni 86212 cB4Documento6 pagineSadava Soluzioni 86212 cB4xavaf31739Nessuna valutazione finora
- Antireumatici 1Documento35 pagineAntireumatici 1bitterlemon01Nessuna valutazione finora
- E EMME Bevanda Sygnum PDFDocumento5 pagineE EMME Bevanda Sygnum PDFHssNessuna valutazione finora
- Batteri Gruppo 1Documento42 pagineBatteri Gruppo 1Domenico RicciardiNessuna valutazione finora
- Miv SuperantigeniDocumento1 paginaMiv Superantigeniantonio nennaNessuna valutazione finora
- Locci 2Documento7 pagineLocci 2Nicola FattoriniNessuna valutazione finora
- Vescicole EndolisosomialiDocumento3 pagineVescicole EndolisosomialiFrancesco BorealeNessuna valutazione finora
- Appunti Di BatteriologiaDocumento46 pagineAppunti Di BatteriologiagabbolNessuna valutazione finora
- Patologia Generale La FebbreDocumento3 paginePatologia Generale La FebbreYassine MandarisNessuna valutazione finora
- Microbiologia e Virologia ClinicaDocumento12 pagineMicrobiologia e Virologia ClinicaokaiusvestaNessuna valutazione finora
- Gramneg LEZIONE Rianimazione 2010Documento52 pagineGramneg LEZIONE Rianimazione 2010Agnese ValentiniNessuna valutazione finora
- Morte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cDa EverandMorte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cNessuna valutazione finora
- Encefalopatia EpaticaDocumento2 pagineEncefalopatia EpaticaGilbertoBruciaferriNessuna valutazione finora
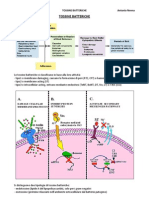
- BatteriologiaDocumento44 pagineBatteriologiaFrancesco D'UrbanoNessuna valutazione finora
- Baicalina SR ITADocumento4 pagineBaicalina SR ITAGiaNessuna valutazione finora
- Coagulaszione Fase Vascolare-PiastrinicaDocumento34 pagineCoagulaszione Fase Vascolare-PiastrinicaRita ContiNessuna valutazione finora
- Lezione 4 - Patologia Genetica 2Documento51 pagineLezione 4 - Patologia Genetica 2lucaNessuna valutazione finora
- 04 SporaDocumento26 pagine04 Sporadschill2709Nessuna valutazione finora
- 2102 Classificazione-AntibioticiDocumento10 pagine2102 Classificazione-AntibioticiteoNessuna valutazione finora
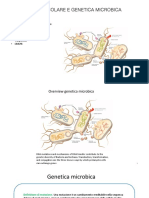
- Genetica Microbica 1 Trasformazione-Trasduzione PDFDocumento33 pagineGenetica Microbica 1 Trasformazione-Trasduzione PDFFederica ZanellaNessuna valutazione finora
- I.2. Otite Acute Si CroniceDocumento43 pagineI.2. Otite Acute Si CroniceMihaela MikiNessuna valutazione finora
- La PlacaDocumento47 pagineLa PlacaFabrizio BarbagalloNessuna valutazione finora
- Sinergismo2 Antinfettivo e AntitumoraleDocumento26 pagineSinergismo2 Antinfettivo e AntitumoraleJulien JulienNessuna valutazione finora
- FansDocumento70 pagineFansParide Di StasioNessuna valutazione finora
- BotulismoDocumento23 pagineBotulismoSilvia PNessuna valutazione finora
- Leucemia Acuta Linfoide PDFDocumento4 pagineLeucemia Acuta Linfoide PDFFrancesco ScognamiglioNessuna valutazione finora
- 6 Enterobacteriaceae 120423122540 Phpapp02Documento33 pagine6 Enterobacteriaceae 120423122540 Phpapp02agostinoburla4446Nessuna valutazione finora
- Lezione 4 (19 Ottobre)Documento7 pagineLezione 4 (19 Ottobre)ste MassariNessuna valutazione finora
- Battaglioli BiologiaCellulare Lez3Documento67 pagineBattaglioli BiologiaCellulare Lez3Renata WürziusNessuna valutazione finora
- Explorare Hepato BiliaraDocumento3 pagineExplorare Hepato BiliaraMirela Sfaraiala LuculescuNessuna valutazione finora
- Miv Sistemi Di SecrezioneDocumento11 pagineMiv Sistemi Di Secrezioneantonio nennaNessuna valutazione finora
- F00b-Controllo EpigeneticoDocumento20 pagineF00b-Controllo Epigeneticoantonio nennaNessuna valutazione finora
- Esotossine Ed EndotossineDocumento3 pagineEsotossine Ed EndotossineLauraNessuna valutazione finora
- Microbiologia Di FiliceDocumento160 pagineMicrobiologia Di FiliceErvis AliajNessuna valutazione finora
- 20181010-Patologia Genetica RGDocumento51 pagine20181010-Patologia Genetica RGlucaNessuna valutazione finora
- Biologia 41 - Regolazione Dell'espressione GenicaDocumento4 pagineBiologia 41 - Regolazione Dell'espressione GenicaFrancesco TarulloNessuna valutazione finora
- 1415 Bbg-Ii Coli 4 PPDFDocumento42 pagine1415 Bbg-Ii Coli 4 PPDFfrancescamariacNessuna valutazione finora
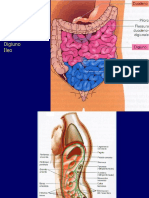
- 2021 Farmacia Digerente 2 23 26 AprileDocumento78 pagine2021 Farmacia Digerente 2 23 26 AprileChiara ArdauNessuna valutazione finora
- Schema Riassuntivo BatteriDocumento55 pagineSchema Riassuntivo BatterifraNessuna valutazione finora
- Omeo StasiDocumento41 pagineOmeo StasiJessica Raffaella MaderaNessuna valutazione finora
- F00e AterosclerosiDocumento10 pagineF00e Aterosclerosiantonio nennaNessuna valutazione finora
- 10 Sanguinamento DigestivoDocumento7 pagine10 Sanguinamento Digestivoantonio nennaNessuna valutazione finora
- F00f-Oncologia EpidemiologiaDocumento6 pagineF00f-Oncologia Epidemiologiaantonio nennaNessuna valutazione finora
- F00c Patologie EpigeneticheDocumento9 pagineF00c Patologie Epigeneticheantonio nenna100% (1)
- F00b-Controllo EpigeneticoDocumento20 pagineF00b-Controllo Epigeneticoantonio nennaNessuna valutazione finora
- F00d Metabolismo LipoproteicoDocumento7 pagineF00d Metabolismo Lipoproteicoantonio nennaNessuna valutazione finora
- F00a-Individualità GeneticaDocumento16 pagineF00a-Individualità Geneticaantonio nennaNessuna valutazione finora
- Chirurgica 1° SemDocumento4 pagineChirurgica 1° Semantonio nennaNessuna valutazione finora
- 13 Equilibrio Acido-BaseDocumento6 pagine13 Equilibrio Acido-Baseantonio nennaNessuna valutazione finora
- Medica 1° SemDocumento4 pagineMedica 1° Semantonio nennaNessuna valutazione finora
- 11 EmostasiDocumento4 pagine11 Emostasiantonio nennaNessuna valutazione finora
- 12 IponatremiaDocumento6 pagine12 Iponatremiaantonio nennaNessuna valutazione finora
- Miv Esperimenti StoriciDocumento8 pagineMiv Esperimenti Storiciantonio nennaNessuna valutazione finora
- Miv ImmunologiaDocumento24 pagineMiv Immunologiaantonio nennaNessuna valutazione finora
- Miv Divisione BattericaDocumento12 pagineMiv Divisione Battericaantonio nennaNessuna valutazione finora
- Miv Struttura Dei BatteriDocumento7 pagineMiv Struttura Dei Batteriantonio nennaNessuna valutazione finora
- Miv Principali Batteri PatogeniDocumento38 pagineMiv Principali Batteri Patogeniantonio nennaNessuna valutazione finora
- Miv Sistemi Di SecrezioneDocumento11 pagineMiv Sistemi Di Secrezioneantonio nennaNessuna valutazione finora
- Miv Replicazione Dei PlasmidiDocumento2 pagineMiv Replicazione Dei Plasmidiantonio nennaNessuna valutazione finora
- Miv Tossine BattericheDocumento6 pagineMiv Tossine Battericheantonio nennaNessuna valutazione finora
- Miv Treponema PallidumDocumento2 pagineMiv Treponema Pallidumantonio nennaNessuna valutazione finora
- Miv Replicazione Dei PlasmidiDocumento2 pagineMiv Replicazione Dei Plasmidiantonio nennaNessuna valutazione finora
- Miv AntibioticiDocumento5 pagineMiv Antibioticiantonio nennaNessuna valutazione finora
- Miv SuperantigeniDocumento1 paginaMiv Superantigeniantonio nennaNessuna valutazione finora
- Miv Tossine BattericheDocumento18 pagineMiv Tossine Battericheantonio nennaNessuna valutazione finora
- Miv Sistemi Di SecrezioneDocumento5 pagineMiv Sistemi Di Secrezioneantonio nennaNessuna valutazione finora
- 8 Vettori e ClonaggioDocumento6 pagine8 Vettori e ClonaggiobvincenzoNessuna valutazione finora
- Bio Chi MicaDocumento242 pagineBio Chi Micalagapij262Nessuna valutazione finora
- 07010000-PI Biotech ItaliaDocumento68 pagine07010000-PI Biotech Italiapedro41Nessuna valutazione finora
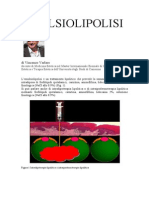
- Varlaro EmulsiolipolisiDocumento37 pagineVarlaro EmulsiolipolisiAngela Pagliuso100% (1)
- La CellulaDocumento2 pagineLa CellulaVittorio AnginiNessuna valutazione finora
- Compendio Di Biologia GeneticaDocumento72 pagineCompendio Di Biologia GeneticaEugenia DantonioNessuna valutazione finora
- Ossidazione Della Cisteina - Scienza e Tecnologia - Notizie Di Chimica e IngegneriaDocumento3 pagineOssidazione Della Cisteina - Scienza e Tecnologia - Notizie Di Chimica e IngegneriaChiara VaccaNessuna valutazione finora
- Sadava Nuova Biologiablu Sintesi A6Documento2 pagineSadava Nuova Biologiablu Sintesi A6Franco 06Nessuna valutazione finora
- VirologiaDocumento16 pagineVirologiasweet_05Nessuna valutazione finora
- 01 Wine BookDocumento100 pagine01 Wine BookИрина ГорбачеваNessuna valutazione finora
- Diabete MelitoDocumento29 pagineDiabete MelitoElton Tony KazaziNessuna valutazione finora