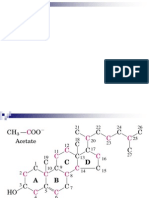
Potrebbero piacerti anche
- Insulina 2018Documento56 pagineInsulina 2018Filippo NovelliNessuna valutazione finora
- GlicolisiDocumento43 pagineGlicolisigaiamgmalerbaNessuna valutazione finora
- Biochimica II - DomandeDocumento15 pagineBiochimica II - DomandeLorenzo Maria CerviNessuna valutazione finora
- GlicolisiDocumento25 pagineGlicolisioliser671Nessuna valutazione finora
- EserciziDocumento26 pagineEsercizidarkgianluca18Nessuna valutazione finora
- ILMETABOLISMOENERGETICODocumento23 pagineILMETABOLISMOENERGETICOdavide abbattistaNessuna valutazione finora
- 3 GlicolisiDocumento48 pagine3 GlicolisiRoberta Lo IzzoNessuna valutazione finora
- GlicolisiDocumento2 pagineGlicolisiCristian UsbergoNessuna valutazione finora
- BiochimicaDocumento29 pagineBiochimicaRoberta PirasNessuna valutazione finora
- 3-Metabolismo Dei GlucidiDocumento39 pagine3-Metabolismo Dei GlucidiFrancesco Iavarone100% (1)
- GlicogenesiDocumento14 pagineGlicogenesiciccioNessuna valutazione finora
- Schemi BiochimicaDocumento14 pagineSchemi Biochimicachucrofr100% (2)
- CicloDigiuno AlimentazioneDocumento44 pagineCicloDigiuno AlimentazioneChiara LassoniNessuna valutazione finora
- 17 Via Dei Pentoso FosfatiDocumento28 pagine17 Via Dei Pentoso Fosfatigabries27Nessuna valutazione finora
- Il Metabolismo Energetico, Sintesi Del Corso Di Scienze Della TerraDocumento9 pagineIl Metabolismo Energetico, Sintesi Del Corso Di Scienze Della TerraluludevergottiniNessuna valutazione finora
- Uomo e NaturaDocumento6 pagineUomo e NaturaAlessia PetragalloNessuna valutazione finora
- I CarboidratiDocumento13 pagineI CarboidratimizarNessuna valutazione finora
- Appunti MetabolismoDocumento19 pagineAppunti MetabolismoLuca CanegratiNessuna valutazione finora
- Biochimica Del FegatoDocumento6 pagineBiochimica Del FegatoSalò MarzialiNessuna valutazione finora
- Metabolismo EnergeticoDocumento4 pagineMetabolismo EnergeticoHghh NgthhNessuna valutazione finora
- Il Metabolismo Dei CarboidratiDocumento7 pagineIl Metabolismo Dei CarboidratiArantxa HoyleNessuna valutazione finora
- Risposte A Domande BiochimicaDocumento7 pagineRisposte A Domande BiochimicaLuigia CentolaNessuna valutazione finora
- GottaDocumento9 pagineGottaannaricci.aNessuna valutazione finora
- Caso Clinico 2Documento8 pagineCaso Clinico 2CarlosFernanGutierrezNessuna valutazione finora
- Metabolismo Delle ProteineDocumento10 pagineMetabolismo Delle ProteineGianluca Garozzo100% (1)
- 10 Gruppo SlidesDocumento18 pagine10 Gruppo SlidesMariacarmela MorrealeNessuna valutazione finora
- Metabolismo CellulareDocumento5 pagineMetabolismo CellulareLeonardo TovoNessuna valutazione finora
- Biochimica Esame 2Documento13 pagineBiochimica Esame 2enzocavajuoloNessuna valutazione finora
- Dommande Con Risposta BiochimicaDocumento37 pagineDommande Con Risposta BiochimicayuppisadNessuna valutazione finora
- Il MetabolismoDocumento5 pagineIl MetabolismoAlice PizziniNessuna valutazione finora
- Ciclo Glucosio AlaninaDocumento2 pagineCiclo Glucosio AlaninaRiccardo ChecchinNessuna valutazione finora
- Lezione 9Documento54 pagineLezione 9Tonia PreziusoNessuna valutazione finora
- Respirazione e Fermentazione-MicrobiologiaDocumento18 pagineRespirazione e Fermentazione-MicrobiologiaSamuel Di sottoNessuna valutazione finora
- Metabolismo MicrobicoDocumento19 pagineMetabolismo Microbicodschill2709100% (3)
- BIOCHIMICA - SiliprandiDocumento50 pagineBIOCHIMICA - SiliprandiinfernuxNessuna valutazione finora
- Metabolismo CellulareDocumento4 pagineMetabolismo CellulareGiada Di GennaroNessuna valutazione finora
- 4 Crisis Hiperglicemia Del AdultoDocumento12 pagine4 Crisis Hiperglicemia Del AdultoKARLA VASQUEZNessuna valutazione finora
- A1.2d - Farmaci Antiperlipemici (4 - 12-2019)Documento64 pagineA1.2d - Farmaci Antiperlipemici (4 - 12-2019)Massimo PregnolatoNessuna valutazione finora
- Lezione 17 Fine DiabeteDocumento27 pagineLezione 17 Fine DiabeteChiara PaolicelliNessuna valutazione finora
- Insulina e GHDocumento8 pagineInsulina e GHNunzia LeoneNessuna valutazione finora
- Ho Lester OlDocumento49 pagineHo Lester OlRoki Popuslic MaliNessuna valutazione finora
- ProstaglandineDocumento26 pagineProstaglandineCezara ZahariaNessuna valutazione finora
- Biologia GeneraleDocumento4 pagineBiologia GeneraleTereresa2Nessuna valutazione finora
- Metabolismo Anaerobico Durante Circolazione ExtracorpDocumento46 pagineMetabolismo Anaerobico Durante Circolazione ExtracorpEffetto Di ProssimitàNessuna valutazione finora
- BioenergeticaDocumento49 pagineBioenergeticaSofia AlvanoNessuna valutazione finora
- Fisiologia e Fisiopatologia Del Diabete MellitoDocumento10 pagineFisiologia e Fisiopatologia Del Diabete Mellitosiluma11100% (1)
- L'insulinaDocumento5 pagineL'insulinavmelani276Nessuna valutazione finora
- A Metabolismo CellulareDocumento14 pagineA Metabolismo CellularemodenafraNessuna valutazione finora
- LEZIONE 9 Ossidazione Acidi GrassiDocumento58 pagineLEZIONE 9 Ossidazione Acidi GrassiMaegazine EnizageamNessuna valutazione finora
- Domande BiochimDocumento11 pagineDomande BiochimAlanNessuna valutazione finora
- Glicolisi PDFDocumento9 pagineGlicolisi PDFSpennacchioneNessuna valutazione finora
- Nozioni NutrizioneDocumento13 pagineNozioni NutrizionePietro PensoNessuna valutazione finora
- Respirazione CellulareDocumento25 pagineRespirazione CellulareLuca GallitelliNessuna valutazione finora
- Biochimica - Scienze Motorie PDFDocumento282 pagineBiochimica - Scienze Motorie PDFRenatoNessuna valutazione finora
- BiochimicDocumento282 pagineBiochimicSalvatore FontanarosaNessuna valutazione finora
- Very Low CHO Keto DietDocumento11 pagineVery Low CHO Keto DietantoniozanniniNessuna valutazione finora
- Ketogenic Diet for Beginners: The Ultimate Keto Diet Guide For Healing Your Body And Aiding Weight Loss (With Over 40 Delicious Recipes)Da EverandKetogenic Diet for Beginners: The Ultimate Keto Diet Guide For Healing Your Body And Aiding Weight Loss (With Over 40 Delicious Recipes)Nessuna valutazione finora
- Dieta chetogenica per principianti: L'ultima guida alla dieta Keto per guarire il tuo corpo e aiutare la perdita di peso (Con oltre 40 deliziose ricette)Da EverandDieta chetogenica per principianti: L'ultima guida alla dieta Keto per guarire il tuo corpo e aiutare la perdita di peso (Con oltre 40 deliziose ricette)Nessuna valutazione finora
- EFFETTI COGNITIVI E FISIOLOGICI DELL' ASSUNZIONE DI ACIDI GRASSI POLINSATURI OMEGA-3 IN SOGGETTI SANIDa EverandEFFETTI COGNITIVI E FISIOLOGICI DELL' ASSUNZIONE DI ACIDI GRASSI POLINSATURI OMEGA-3 IN SOGGETTI SANINessuna valutazione finora
- 5 - La ForzaDocumento50 pagine5 - La ForzaMaegazine EnizageamNessuna valutazione finora
- 2 - L'attività SportivaDocumento24 pagine2 - L'attività SportivaMaegazine EnizageamNessuna valutazione finora
- 4 - La TecnicaDocumento41 pagine4 - La TecnicaMaegazine EnizageamNessuna valutazione finora
- 3 - Le Capacità CordinativeDocumento39 pagine3 - Le Capacità CordinativeMaegazine EnizageamNessuna valutazione finora
- TTD - TMU - Lezioni I - XIIDocumento81 pagineTTD - TMU - Lezioni I - XIIMaegazine EnizageamNessuna valutazione finora
- TTD - TMU - Lezioni I - II - IIIDocumento17 pagineTTD - TMU - Lezioni I - II - IIIMaegazine EnizageamNessuna valutazione finora
- Lezioni I e IIDocumento15 pagineLezioni I e IIMaegazine EnizageamNessuna valutazione finora
- Letteratura TTD Attività Motorie e SportiveDocumento1 paginaLetteratura TTD Attività Motorie e SportiveMaegazine EnizageamNessuna valutazione finora
- Dispensa ScienzemotorieDocumento72 pagineDispensa ScienzemotorieMaegazine EnizageamNessuna valutazione finora
- Lez1 - PresentazioneDocumento32 pagineLez1 - PresentazioneMaegazine EnizageamNessuna valutazione finora
- I Lettori Rispondono A RousseauDocumento1 paginaI Lettori Rispondono A RousseauMaegazine EnizageamNessuna valutazione finora
- Linguistica RomanzaDocumento27 pagineLinguistica RomanzaMaegazine EnizageamNessuna valutazione finora
- Schema 1Documento3 pagineSchema 1Maegazine EnizageamNessuna valutazione finora
- 06intro Metabolismo MicrobicoDocumento53 pagine06intro Metabolismo Microbicodschill2709100% (1)
- Respirazione CellulareDocumento25 pagineRespirazione CellulareLuca GallitelliNessuna valutazione finora
- Fisiologia UmanaDocumento254 pagineFisiologia UmanagigioNessuna valutazione finora
- CITOLOGIA E ISTOLOGIA - Lezione 12Documento4 pagineCITOLOGIA E ISTOLOGIA - Lezione 12Federico TosiniNessuna valutazione finora
- Apparato Circolatorio PDFDocumento26 pagineApparato Circolatorio PDFSand TasticNessuna valutazione finora
- Protocolli EPOCDocumento1 paginaProtocolli EPOCAndrea Dell'angeloNessuna valutazione finora
- 2019-12-17 Fisiologia 1 - Lezione 44Documento10 pagine2019-12-17 Fisiologia 1 - Lezione 44Lucia SparmajerNessuna valutazione finora