Potrebbero piacerti anche
- Nanotecnologie per il trattamento dei tumori: quali sono gli effetti sul genoma?Da EverandNanotecnologie per il trattamento dei tumori: quali sono gli effetti sul genoma?Nessuna valutazione finora
- Osteopatia Essenziale e tecniche sull'area lombo-pelvicaDa EverandOsteopatia Essenziale e tecniche sull'area lombo-pelvicaNessuna valutazione finora
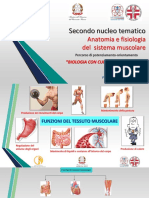
- Fisiologia Del MuscoloDocumento44 pagineFisiologia Del MuscoloGiacomo SaiuNessuna valutazione finora
- 5 Contrazione MuscolareDocumento26 pagine5 Contrazione MuscolareLuisa GrossoNessuna valutazione finora
- Muscolo Schel - Parte1Documento34 pagineMuscolo Schel - Parte1Francesco MalavoltaNessuna valutazione finora
- Il MuscoloDocumento25 pagineIl MuscoloSimone CipollettiNessuna valutazione finora
- Contrazione MuscolareDocumento15 pagineContrazione Muscolareapi-26155364100% (4)
- Apparato MuscolareDocumento4 pagineApparato MuscolareMatteo GrassoNessuna valutazione finora
- 16 - Contrazione Muscolare PDFDocumento32 pagine16 - Contrazione Muscolare PDFcipNessuna valutazione finora
- Biochimica Del MuscoloDocumento32 pagineBiochimica Del MuscoloAntonio IaconetaNessuna valutazione finora
- AAA Elettroterapia PDFDocumento51 pagineAAA Elettroterapia PDFOsteofisioNessuna valutazione finora
- Anatomia e Fisiologia Del Sistema MuscolareDocumento41 pagineAnatomia e Fisiologia Del Sistema MuscolareHelloNessuna valutazione finora
- 1 - Introduzione e MembranaDocumento28 pagine1 - Introduzione e Membranaapi-26155364Nessuna valutazione finora
- Fisiologia Del Muscolo ScheletricoDocumento20 pagineFisiologia Del Muscolo ScheletricoCarlo BritoNessuna valutazione finora
- 7-8 Tessuto e Sistema MuscolareDocumento9 pagine7-8 Tessuto e Sistema Muscolaremariannaguadagno00Nessuna valutazione finora
- Accoppiamento Eccitazione-Contrazione Nel CuoreDocumento35 pagineAccoppiamento Eccitazione-Contrazione Nel CuoreLalla BozziNessuna valutazione finora
- Scienze 4 VerificaDocumento23 pagineScienze 4 VerificaAlessandra SilveraNessuna valutazione finora
- Proteine MuscolariDocumento18 pagineProteine MuscolariAndrea SanzoNessuna valutazione finora
- 2.fisiologia MuscolareDocumento17 pagine2.fisiologia MuscolareHussam SmithNessuna valutazione finora
- EvoliDocumento93 pagineEvoliPietro BracagliaNessuna valutazione finora
- 01 Muscolo ScheletricoDocumento43 pagine01 Muscolo ScheletricoaxelshippoNessuna valutazione finora
- Istologia 42 - Muscolare Liscio PDFDocumento3 pagineIstologia 42 - Muscolare Liscio PDFAli S SaadNessuna valutazione finora
- 6S - Apparato LocomotoreDocumento7 pagine6S - Apparato LocomotoreAlessandra FrassoNessuna valutazione finora
- Tessuto Muscolare MedicinaDocumento24 pagineTessuto Muscolare MedicinaAlessandro MonteNessuna valutazione finora
- Anatomia e Fisiologia Del Sistema MuscolareDocumento36 pagineAnatomia e Fisiologia Del Sistema MuscolareFrancescaNessuna valutazione finora
- Il Tessuto MuscolareDocumento19 pagineIl Tessuto MuscolareAnnamaria MaderaNessuna valutazione finora
- TRAINER Liv 1Documento62 pagineTRAINER Liv 1Paolo GalbiatiNessuna valutazione finora
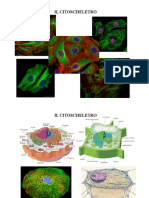
- Citoscheletro e Motilità Cellulare - Capitolo 7Documento4 pagineCitoscheletro e Motilità Cellulare - Capitolo 7maiana91Nessuna valutazione finora
- Tessuto MuscolareDocumento77 pagineTessuto Muscolareilaria zegaNessuna valutazione finora
- MITOCONDRIDocumento4 pagineMITOCONDRIMartina PorcuNessuna valutazione finora
- Muscolo ScheletricoDocumento4 pagineMuscolo ScheletricoHghh NgthhNessuna valutazione finora
- RIASSUNTO MUSCOLI (Cap.12-14-15)Documento8 pagineRIASSUNTO MUSCOLI (Cap.12-14-15)ANNALISA ZINNINessuna valutazione finora
- Biologia TessutiDocumento5 pagineBiologia TessutiFederica CadoniNessuna valutazione finora
- Fisiologia Dell' Esercizio e Dello SportDocumento65 pagineFisiologia Dell' Esercizio e Dello SportDaniele Nervi33% (3)
- Tessuto Muscolare PDFDocumento15 pagineTessuto Muscolare PDFLuis CriscuoloNessuna valutazione finora
- Muscolare Gen Scheletrico L16b (10 11)Documento16 pagineMuscolare Gen Scheletrico L16b (10 11)Lalla BozziNessuna valutazione finora
- Tessuto MuscolareDocumento3 pagineTessuto Muscolaredulli91Nessuna valutazione finora
- 2 Riassunto CitologiaDocumento14 pagine2 Riassunto CitologiaSilvia ZottiNessuna valutazione finora
- CitoscheletroDocumento43 pagineCitoscheletromartinaNessuna valutazione finora
- 9 Eucariote Animale - Citoscheletro, CentrioliDocumento8 pagine9 Eucariote Animale - Citoscheletro, CentrioliLuana LeonardiNessuna valutazione finora
- Apparato MuscolareDocumento14 pagineApparato MuscolaremelissaNessuna valutazione finora
- Tessuto NervosoDocumento6 pagineTessuto NervosoDinjo79Nessuna valutazione finora
- Muscolo LiscioDocumento14 pagineMuscolo LiscioDaniela PiteoNessuna valutazione finora
- La Programmazione Degli AllenamentiDocumento44 pagineLa Programmazione Degli Allenamentifilippo.2003.f8Nessuna valutazione finora
- Il Sistema Muscolo-ScheletricoDocumento4 pagineIl Sistema Muscolo-ScheletricoCarlotta MazzoniNessuna valutazione finora
- Fisiología 7Documento9 pagineFisiología 7mrvbn8tcvwNessuna valutazione finora
- Apparato MuscolareDocumento18 pagineApparato MuscolarecNessuna valutazione finora
- C-Muscoli BiotecDocumento32 pagineC-Muscoli BiotecLalla BozziNessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- Meccanica MuscolareDocumento25 pagineMeccanica Muscolareleo.sei.beloNessuna valutazione finora
- CITOSCHELETRODocumento3 pagineCITOSCHELETROLisaNessuna valutazione finora
- Release Miofasciale Ente Italia SportDocumento56 pagineRelease Miofasciale Ente Italia Sportwnsjt7gdpzNessuna valutazione finora
- Potenziale D'azione CardiacoDocumento53 paginePotenziale D'azione CardiacoLalla BozziNessuna valutazione finora
- I MUSCOLI RiassuntoDocumento6 pagineI MUSCOLI RiassuntoFrancesca MarconeNessuna valutazione finora
- Appunti Settimana 1 CBI ITADocumento10 pagineAppunti Settimana 1 CBI ITALuna SartoriNessuna valutazione finora
- Il MitocondrioDocumento40 pagineIl Mitocondriozeeus2013Nessuna valutazione finora
- Muscolo LiscioDocumento20 pagineMuscolo Liscioleo.sei.beloNessuna valutazione finora
- Tessuto Muscolare: Il Tessuto Muscolare È Responsabile Dei Movimenti Volontari Ed Involontari Del CorpoDocumento4 pagineTessuto Muscolare: Il Tessuto Muscolare È Responsabile Dei Movimenti Volontari Ed Involontari Del CorpoLuana LeonardiNessuna valutazione finora
- CitoscheletroDocumento11 pagineCitoscheletroCris PecoraNessuna valutazione finora
- DEACETILASI ISTONICHE QUALI POTENZIALI BERSAGLI FARMACEUTICIDa EverandDEACETILASI ISTONICHE QUALI POTENZIALI BERSAGLI FARMACEUTICINessuna valutazione finora
- V Lez - APA - 2018 - Malattie Croniche e PS PDFDocumento52 pagineV Lez - APA - 2018 - Malattie Croniche e PS PDFXXXXXXXXNessuna valutazione finora
- TTD Motoria - MONACI 30-10-2019Documento3 pagineTTD Motoria - MONACI 30-10-2019XXXXXXXXNessuna valutazione finora
- V Lez - APA - 2018 - Malattie Croniche e PS PDFDocumento52 pagineV Lez - APA - 2018 - Malattie Croniche e PS PDFXXXXXXXXNessuna valutazione finora
- Traumi Da SportDocumento32 pagineTraumi Da SportXXXXXXXXNessuna valutazione finora
- Laurea Da CaricareDocumento72 pagineLaurea Da CaricareXXXXXXXXNessuna valutazione finora
- TTD Motoria 14Documento7 pagineTTD Motoria 14XXXXXXXXNessuna valutazione finora
- ULTIMO AGGIORAMETO Ortogr Pag.7Documento76 pagineULTIMO AGGIORAMETO Ortogr Pag.7XXXXXXXXNessuna valutazione finora
- VERCELLI Lez 3 2019 PDFDocumento25 pagineVERCELLI Lez 3 2019 PDFXXXXXXXXNessuna valutazione finora
- ULTIMO AGGIORAMETO Ortogr Pag.7Documento76 pagineULTIMO AGGIORAMETO Ortogr Pag.7XXXXXXXXNessuna valutazione finora
- TTD Motoria 14Documento7 pagineTTD Motoria 14XXXXXXXXNessuna valutazione finora
- VERCELLI Lez 4 2019 PDFDocumento44 pagineVERCELLI Lez 4 2019 PDFXXXXXXXXNessuna valutazione finora
- Traumi Da SportDocumento32 pagineTraumi Da SportXXXXXXXXNessuna valutazione finora
- VERCELLI Lez 1-2 2019 PDFDocumento64 pagineVERCELLI Lez 1-2 2019 PDFXXXXXXXXNessuna valutazione finora
- Laurea Da CaricareDocumento72 pagineLaurea Da CaricareXXXXXXXXNessuna valutazione finora
- Ricerca Cardio e VolemiaDocumento3 pagineRicerca Cardio e VolemiaXXXXXXXXNessuna valutazione finora
- TTD Motoria - MONACI 30-10-2019Documento3 pagineTTD Motoria - MONACI 30-10-2019XXXXXXXXNessuna valutazione finora
- 3 Eserciziocardiovasc 2019 PDFDocumento29 pagine3 Eserciziocardiovasc 2019 PDFXXXXXXXXNessuna valutazione finora
- 2 Regolazionepressione 2019 PDFDocumento23 pagine2 Regolazionepressione 2019 PDFXXXXXXXXNessuna valutazione finora
- L Sistema ReninaDocumento2 pagineL Sistema ReninaXXXXXXXXNessuna valutazione finora
- Ultima CappelliniDocumento3 pagineUltima CappelliniXXXXXXXXNessuna valutazione finora
- 1 Cardiocircolatorio 2019 PDFDocumento16 pagine1 Cardiocircolatorio 2019 PDFXXXXXXXXNessuna valutazione finora
- Peptidi NatriureticiDocumento1 paginaPeptidi NatriureticiXXXXXXXXNessuna valutazione finora
- VOLEMIADocumento2 pagineVOLEMIAXXXXXXXXNessuna valutazione finora
- Muscolo ScheletricoDocumento2 pagineMuscolo ScheletricoXXXXXXXXNessuna valutazione finora
- Adattamenti Respiratori IesercizioDocumento3 pagineAdattamenti Respiratori IesercizioXXXXXXXXNessuna valutazione finora
- 01 Pellegrini Intro PDFDocumento47 pagine01 Pellegrini Intro PDFXXXXXXXXNessuna valutazione finora
- Adattamenti All Esercizio FisicoDocumento3 pagineAdattamenti All Esercizio FisicoXXXXXXXXNessuna valutazione finora
- NUTRIZIONEDocumento4 pagineNUTRIZIONEXXXXXXXXNessuna valutazione finora
- Ciclo CoronaricoDocumento2 pagineCiclo CoronaricoXXXXXXXXNessuna valutazione finora
- 03 Pellegrini - AntidepressiviDocumento29 pagine03 Pellegrini - AntidepressiviXXXXXXXXNessuna valutazione finora
- Lezione Di AnatomiaDocumento9 pagineLezione Di AnatomiaMarinela GiciuNessuna valutazione finora
- Ch07 LaPercezioneDelMovimentoDocumento14 pagineCh07 LaPercezioneDelMovimentoAlfonso BelfioreNessuna valutazione finora
- Chimica Delle FermentazioniDocumento87 pagineChimica Delle FermentazionimichNessuna valutazione finora
- Magnani Riscaldamento GiovanileDocumento88 pagineMagnani Riscaldamento GiovanileMohamed JelassiNessuna valutazione finora
- Ocluzia Intestinala LicentaDocumento147 pagineOcluzia Intestinala LicentaVladutz GeorgianNessuna valutazione finora