Potrebbero piacerti anche
- Esercizi di fisica: fisica nucleare e fisica della materiaDa EverandEsercizi di fisica: fisica nucleare e fisica della materiaNessuna valutazione finora
- 1 - Introduzione e MembranaDocumento28 pagine1 - Introduzione e Membranaapi-26155364Nessuna valutazione finora
- Lezione 1 Contrazione Muscolare PDFDocumento75 pagineLezione 1 Contrazione Muscolare PDFXXXXXXXXNessuna valutazione finora
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- Muscolo Schel - Parte1Documento34 pagineMuscolo Schel - Parte1Francesco MalavoltaNessuna valutazione finora
- Nanotecnologie per il trattamento dei tumori: quali sono gli effetti sul genoma?Da EverandNanotecnologie per il trattamento dei tumori: quali sono gli effetti sul genoma?Nessuna valutazione finora
- Fisiologia Del MuscoloDocumento44 pagineFisiologia Del MuscoloGiacomo SaiuNessuna valutazione finora
- Esseri Viventi Dipendono Dalla Meccanica QuantisticaDa EverandEsseri Viventi Dipendono Dalla Meccanica QuantisticaNessuna valutazione finora
- 5 Contrazione MuscolareDocumento26 pagine5 Contrazione MuscolareLuisa GrossoNessuna valutazione finora
- Il MuscoloDocumento25 pagineIl MuscoloSimone CipollettiNessuna valutazione finora
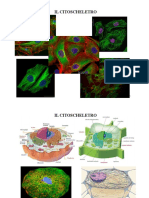
- CitoscheletroDocumento43 pagineCitoscheletromartinaNessuna valutazione finora
- Apparato MuscolareDocumento4 pagineApparato MuscolareMatteo GrassoNessuna valutazione finora
- Meccanica MuscolareDocumento73 pagineMeccanica MuscolareGiovanniNessuna valutazione finora
- 2 Riassunto CitologiaDocumento14 pagine2 Riassunto CitologiaSilvia ZottiNessuna valutazione finora
- 16 - Contrazione Muscolare PDFDocumento32 pagine16 - Contrazione Muscolare PDFcipNessuna valutazione finora
- Fisiologia Del Muscolo ScheletricoDocumento20 pagineFisiologia Del Muscolo ScheletricoCarlo BritoNessuna valutazione finora
- Accoppiamento Eccitazione-Contrazione Nel CuoreDocumento35 pagineAccoppiamento Eccitazione-Contrazione Nel CuoreLalla BozziNessuna valutazione finora
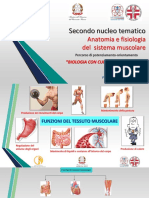
- Anatomia e Fisiologia Del Sistema MuscolareDocumento41 pagineAnatomia e Fisiologia Del Sistema MuscolareHelloNessuna valutazione finora
- CentrioliDocumento26 pagineCentrioliValeria BrancaNessuna valutazione finora
- Meccanica MuscolareDocumento25 pagineMeccanica Muscolareleo.sei.beloNessuna valutazione finora
- RM I Parametri e La SemeioticaDocumento33 pagineRM I Parametri e La SemeioticaIlenia FilaretiNessuna valutazione finora
- Biochimica Del MuscoloDocumento32 pagineBiochimica Del MuscoloAntonio IaconetaNessuna valutazione finora
- BiomeccanicaDocumento30 pagineBiomeccanicaalexander pillacaNessuna valutazione finora
- Citoscheletro e Motilità Cellulare - Capitolo 7Documento4 pagineCitoscheletro e Motilità Cellulare - Capitolo 7maiana91Nessuna valutazione finora
- Fa Cure 2005Documento4 pagineFa Cure 2005Az FatiNessuna valutazione finora
- CitoscheletroDocumento11 pagineCitoscheletroCris PecoraNessuna valutazione finora
- 2.fisiologia MuscolareDocumento17 pagine2.fisiologia MuscolareHussam SmithNessuna valutazione finora
- 6S - Apparato LocomotoreDocumento7 pagine6S - Apparato LocomotoreAlessandra FrassoNessuna valutazione finora
- TRAINER Liv 1Documento62 pagineTRAINER Liv 1Paolo GalbiatiNessuna valutazione finora
- EMGDocumento47 pagineEMGlubas2Nessuna valutazione finora
- ElettrofisiologiaDocumento12 pagineElettrofisiologiaapi-3774792100% (1)
- Slides Le TendinopatieDocumento65 pagineSlides Le TendinopatieSpodyNessuna valutazione finora
- Il Tessuto MuscolareDocumento19 pagineIl Tessuto MuscolareAnnamaria MaderaNessuna valutazione finora
- Trasmissione SinapticaDocumento12 pagineTrasmissione Sinapticaapi-3774792100% (1)
- Stages 2016 FinalDocumento114 pagineStages 2016 FinalElisa Di CarloNessuna valutazione finora
- Corso App 1Documento38 pagineCorso App 1bobbibidibiNessuna valutazione finora
- AAA Elettroterapia PDFDocumento51 pagineAAA Elettroterapia PDFOsteofisioNessuna valutazione finora
- Elettroliti 201203Documento64 pagineElettroliti 201203setcomNessuna valutazione finora
- La Programmazione Degli AllenamentiDocumento44 pagineLa Programmazione Degli Allenamentifilippo.2003.f8Nessuna valutazione finora
- Cap. 5 Principi Di EmodinamicaDocumento40 pagineCap. 5 Principi Di EmodinamicaFrancescaNessuna valutazione finora
- 7 - Neuroni, Integrazione, NeuromodulazioneDocumento12 pagine7 - Neuroni, Integrazione, Neuromodulazioneapi-26155364Nessuna valutazione finora
- Parte II Trattamento Delle Sequenze Apparato-Fasciali: Stecco08.indd 87Documento18 pagineParte II Trattamento Delle Sequenze Apparato-Fasciali: Stecco08.indd 87Dandino SpellmanNessuna valutazione finora
- 16 061216.pptx LockedDocumento23 pagine16 061216.pptx Lockedlocimo3302Nessuna valutazione finora
- 3 - Esame UT - Parte Terza -Trasduttori -35 [Modalità Compatibilità]Documento36 pagine3 - Esame UT - Parte Terza -Trasduttori -35 [Modalità Compatibilità]isacco89Nessuna valutazione finora
- EvoliDocumento93 pagineEvoliPietro BracagliaNessuna valutazione finora
- Dinamica Delle ProteineDocumento6 pagineDinamica Delle ProteineRaffaele VerdeNessuna valutazione finora
- Biomeccanica Del Tessuto MuscolareDocumento34 pagineBiomeccanica Del Tessuto MuscolareduskyroseNessuna valutazione finora
- Principi Fisici Dei Trasduttori - TESINADocumento7 paginePrincipi Fisici Dei Trasduttori - TESINAgokussj3000Nessuna valutazione finora
- Fisiologia Dell' Esercizio e Dello SportDocumento65 pagineFisiologia Dell' Esercizio e Dello SportDaniele Nervi33% (3)
- Biomeccanica Anatomo FunzionaleDocumento257 pagineBiomeccanica Anatomo FunzionaleFlavio VillaNessuna valutazione finora
- Reti Elettriche PDFDocumento99 pagineReti Elettriche PDFivan_terzoNessuna valutazione finora
- Anatomia e Fisiologia Del Sistema MuscolareDocumento36 pagineAnatomia e Fisiologia Del Sistema MuscolareFrancescaNessuna valutazione finora
- Tessuto Muscolare MedicinaDocumento24 pagineTessuto Muscolare MedicinaAlessandro MonteNessuna valutazione finora
- Transistor InterruttoreDocumento8 pagineTransistor InterruttoreClaudiomeneNessuna valutazione finora
- TecarDocumento4 pagineTecarMichele MarengoNessuna valutazione finora
- Scienze 4 VerificaDocumento23 pagineScienze 4 VerificaAlessandra SilveraNessuna valutazione finora
- MITOCONDRIDocumento4 pagineMITOCONDRIMartina PorcuNessuna valutazione finora
- Esercizi Svolti in AulaDocumento25 pagineEsercizi Svolti in AulaFlaviaDiNapoliNessuna valutazione finora
- Chimica InorganicaDocumento42 pagineChimica Inorganicar925tfw6xyNessuna valutazione finora
- 9 - Recettori Sensoriali, Sistemi IntegratiDocumento14 pagine9 - Recettori Sensoriali, Sistemi Integratiapi-26155364Nessuna valutazione finora
- Trasmissione SinapticaDocumento12 pagineTrasmissione Sinapticaapi-3774792100% (1)
- 2 - Eq - ElettrochimDocumento6 pagine2 - Eq - Elettrochimapi-26155364Nessuna valutazione finora
- 7 - Neuroni, Integrazione, NeuromodulazioneDocumento12 pagine7 - Neuroni, Integrazione, Neuromodulazioneapi-26155364Nessuna valutazione finora
- Contrazione MuscolareDocumento15 pagineContrazione Muscolareapi-26155364100% (4)
- ElettrofisiologiaDocumento12 pagineElettrofisiologiaapi-3774792100% (1)
- Fisiologia Parte 4.pdaDocumento14 pagineFisiologia Parte 4.pdaapi-3774792Nessuna valutazione finora
- Lab Biochimica PaoliDocumento28 pagineLab Biochimica Paoliapi-26155364Nessuna valutazione finora
- TUBERCOLOSI Lanciotti 08 OKDocumento8 pagineTUBERCOLOSI Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- ProgrammaDocumento2 pagineProgrammaapi-26155364Nessuna valutazione finora
- Tifo e Paratifo - Lanciotti08Documento8 pagineTifo e Paratifo - Lanciotti08api-26155364Nessuna valutazione finora
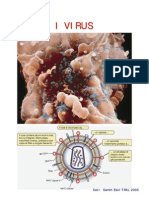
- Virus 08 PPTDocumento26 pagineVirus 08 PPTapi-26155364Nessuna valutazione finora
- 3 - Trasporto AttivoDocumento9 pagine3 - Trasporto Attivoapi-26155364Nessuna valutazione finora
- SPORIGENI&TETANO Lanciotti 08 OKDocumento6 pagineSPORIGENI&TETANO Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- TBC Iss 06Documento4 pagineTBC Iss 06api-26155364Nessuna valutazione finora
- Domande Frequenti PazzagliDocumento1 paginaDomande Frequenti Pazzagliapi-26155364Nessuna valutazione finora
- TBC BonanniDocumento30 pagineTBC Bonanniapi-26155364100% (2)
- RischioBiol LoNostro 06Documento62 pagineRischioBiol LoNostro 06api-26155364Nessuna valutazione finora
- TBC BonanniDocumento30 pagineTBC Bonanniapi-26155364Nessuna valutazione finora
- RischioInf Viaggiatori 08Documento20 pagineRischioInf Viaggiatori 08api-26155364Nessuna valutazione finora
- Poliomielite CrovariDocumento32 paginePoliomielite Crovariapi-26155364Nessuna valutazione finora
- Meningiti ISS 06Documento2 pagineMeningiti ISS 06api-26155364Nessuna valutazione finora
- Influenza CrovariDocumento48 pagineInfluenza Crovariapi-26155364Nessuna valutazione finora
- INFLUENZA Lanciotti 08 OKDocumento47 pagineINFLUENZA Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- INFLUENZA Lanciotti 08 OKDocumento47 pagineINFLUENZA Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- PRIONI&BSE Lanciotti 08 OKDocumento9 paginePRIONI&BSE Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- HB InfermieriDocumento22 pagineHB Infermieriapi-26155364100% (1)
- EpatitiCroniche 06 OKDocumento23 pagineEpatitiCroniche 06 OKapi-26155364Nessuna valutazione finora
- HB InfermieriDocumento22 pagineHB Infermieriapi-26155364100% (1)
- Etologia Del Cavallo PDFDocumento2 pagineEtologia Del Cavallo PDFCarlos0% (1)
- Che Brochure FinalDocumento2 pagineChe Brochure FinalPrincess Janine CatralNessuna valutazione finora
- Anatomia ComparataDocumento106 pagineAnatomia ComparataAndrea GattoNessuna valutazione finora
- Castaldi - Essenze Forestali - Manuale HoepliDocumento435 pagineCastaldi - Essenze Forestali - Manuale HoepliEmanuele MeloniNessuna valutazione finora
- Appunti Di ZoologiaDocumento123 pagineAppunti Di ZoologiaDario Marchese100% (1)








































![3 - Esame UT - Parte Terza -Trasduttori -35 [Modalità Compatibilità]](https://imgv2-1-f.scribdassets.com/img/document/720827198/149x198/e889220999/1712491206?v=1)