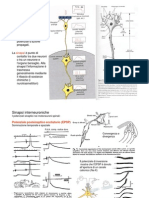
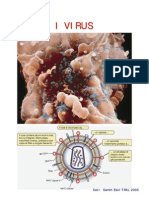
Potrebbero piacerti anche
- Contrazione MuscolareDocumento15 pagineContrazione Muscolareapi-26155364100% (4)
- Fisiologia Parte 4.pdaDocumento14 pagineFisiologia Parte 4.pdaapi-3774792Nessuna valutazione finora
- 2 - Eq - ElettrochimDocumento6 pagine2 - Eq - Elettrochimapi-26155364Nessuna valutazione finora
- 7 - Neuroni, Integrazione, NeuromodulazioneDocumento12 pagine7 - Neuroni, Integrazione, Neuromodulazioneapi-26155364Nessuna valutazione finora
- Trasmissione SinapticaDocumento12 pagineTrasmissione Sinapticaapi-3774792100% (1)
- ElettrofisiologiaDocumento12 pagineElettrofisiologiaapi-3774792100% (1)
- 3 - Trasporto AttivoDocumento9 pagine3 - Trasporto Attivoapi-26155364Nessuna valutazione finora
- Domande Frequenti PazzagliDocumento1 paginaDomande Frequenti Pazzagliapi-26155364Nessuna valutazione finora
- Tifo e Paratifo - Lanciotti08Documento8 pagineTifo e Paratifo - Lanciotti08api-26155364Nessuna valutazione finora
- Virus 08 PPTDocumento26 pagineVirus 08 PPTapi-26155364Nessuna valutazione finora
- TBC Iss 06Documento4 pagineTBC Iss 06api-26155364Nessuna valutazione finora
- TUBERCOLOSI Lanciotti 08 OKDocumento8 pagineTUBERCOLOSI Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- 1 - Introduzione e MembranaDocumento28 pagine1 - Introduzione e Membranaapi-26155364Nessuna valutazione finora
- Lab Biochimica PaoliDocumento28 pagineLab Biochimica Paoliapi-26155364Nessuna valutazione finora
- SPORIGENI&TETANO Lanciotti 08 OKDocumento6 pagineSPORIGENI&TETANO Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- ProgrammaDocumento2 pagineProgrammaapi-26155364Nessuna valutazione finora
- TBC BonanniDocumento30 pagineTBC Bonanniapi-26155364100% (2)
- RischioInf Viaggiatori 08Documento20 pagineRischioInf Viaggiatori 08api-26155364Nessuna valutazione finora
- PRIONI&BSE Lanciotti 08 OKDocumento9 paginePRIONI&BSE Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- TBC BonanniDocumento30 pagineTBC Bonanniapi-26155364Nessuna valutazione finora
- RischioBiol LoNostro 06Documento62 pagineRischioBiol LoNostro 06api-26155364Nessuna valutazione finora
- Meningiti ISS 06Documento2 pagineMeningiti ISS 06api-26155364Nessuna valutazione finora
- INFLUENZA Lanciotti 08 OKDocumento47 pagineINFLUENZA Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- INFLUENZA Lanciotti 08 OKDocumento47 pagineINFLUENZA Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- Influenza CrovariDocumento48 pagineInfluenza Crovariapi-26155364Nessuna valutazione finora
- HB InfermieriDocumento22 pagineHB Infermieriapi-26155364100% (1)
- Poliomielite CrovariDocumento32 paginePoliomielite Crovariapi-26155364Nessuna valutazione finora
- HB InfermieriDocumento22 pagineHB Infermieriapi-26155364100% (1)
- EpatitiCroniche 06 OKDocumento23 pagineEpatitiCroniche 06 OKapi-26155364Nessuna valutazione finora