Potrebbero piacerti anche
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- Guida pratica per ridurre le onde elettromagnetiche: Proteggersi dall’elettrosmog: tutti i rischi per la salute e come difendersiDa EverandGuida pratica per ridurre le onde elettromagnetiche: Proteggersi dall’elettrosmog: tutti i rischi per la salute e come difendersiNessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- Conduzione NervosaDocumento10 pagineConduzione NervosaMiriam LongobardiNessuna valutazione finora
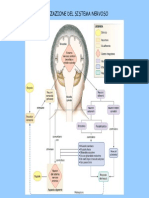
- Il Sistema NervosoDocumento6 pagineIl Sistema NervosoMartina MusmeciNessuna valutazione finora
- FisiologiaDocumento42 pagineFisiologiaLaura TreggiariNessuna valutazione finora
- 01-La Genesi Dell'impulso Nervoso PDFDocumento8 pagine01-La Genesi Dell'impulso Nervoso PDFDiego Flores EgüezNessuna valutazione finora
- Dispensa Fisiologia UnitoDocumento497 pagineDispensa Fisiologia Unitodorian88Nessuna valutazione finora
- FISIOLOGIAsbobineDocumento164 pagineFISIOLOGIAsbobinemarco.ricco.mr00Nessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Fisiologia - Fisica Dell'EcgDocumento23 pagineFisiologia - Fisica Dell'EcgBenjaminMeghnagi0% (1)
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- Fisiopatologia Del Segnale Lezione 1 e 2Documento67 pagineFisiopatologia Del Segnale Lezione 1 e 2Mariagiovanna MariagiovannaNessuna valutazione finora
- Fisiologia PDFDocumento75 pagineFisiologia PDFMarioNessuna valutazione finora
- 2 - BioelettricitàDocumento4 pagine2 - BioelettricitàFabio BordinNessuna valutazione finora
- Potenziali D'azioneDocumento5 paginePotenziali D'azioneHghh NgthhNessuna valutazione finora
- Potenziale Di MembranaDocumento17 paginePotenziale Di MembranamelissaNessuna valutazione finora
- 10 - Potenziali Graduati e Potenziale D-AzioneDocumento48 pagine10 - Potenziali Graduati e Potenziale D-AzioneMarcoNessuna valutazione finora
- CITOLOGIA E ISTOLOGIA - Lezione 7Documento2 pagineCITOLOGIA E ISTOLOGIA - Lezione 7Federico TosiniNessuna valutazione finora
- Segnali BioelettriciDocumento12 pagineSegnali BioelettriciFrancesco BorealeNessuna valutazione finora
- Appunti Di NeurofisiologiaDocumento91 pagineAppunti Di NeurofisiologiaSaraNessuna valutazione finora
- AAA Elettroterapia PDFDocumento51 pagineAAA Elettroterapia PDFOsteofisioNessuna valutazione finora
- 1 Potenziale Di MembranaDocumento2 pagine1 Potenziale Di MembranaGiacomo Ascari-RaccagniNessuna valutazione finora
- 4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azioneDocumento43 pagine4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azionedavide di lorenzoNessuna valutazione finora
- L'impulso NervosoDocumento7 pagineL'impulso NervosoGiancarlo AttivissimoNessuna valutazione finora
- Chimica AnaliticaDocumento2 pagineChimica AnaliticaStefano GioiaNessuna valutazione finora
- I Recettori SensorialiDocumento37 pagineI Recettori SensorialiMaryna100% (1)
- La Pompa Sodio-PotassioDocumento19 pagineLa Pompa Sodio-PotassioDavide ValmorbidaNessuna valutazione finora
- Sbobine Fisiologia Umana I 2015Documento443 pagineSbobine Fisiologia Umana I 2015Leonardo MarchettiNessuna valutazione finora
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- 5 Potenziali ElettrotoniciDocumento8 pagine5 Potenziali ElettrotoniciClaudia OrlannoNessuna valutazione finora
- Docsity Domande Triennale Fisiologia UmanaDocumento33 pagineDocsity Domande Triennale Fisiologia Umanadestinyema78Nessuna valutazione finora
- ElettrocardiogrammaDocumento25 pagineElettrocardiogrammaapi-3704348100% (7)
- Conduzione Impulso NervosoDocumento32 pagineConduzione Impulso NervosoMarcoNessuna valutazione finora
- Fisiologia Veterinaria Completo en Portugues PDFDocumento662 pagineFisiologia Veterinaria Completo en Portugues PDFMartin RpoNessuna valutazione finora
- Riassuntidi BiofisicaDocumento14 pagineRiassuntidi BiofisicaFlavia IovinoNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- NeurofisiologiaDocumento69 pagineNeurofisiologiafabiana900% (1)
- EPRDocumento11 pagineEPR88alexiaNessuna valutazione finora
- Domande PIT FisiologiaDocumento5 pagineDomande PIT FisiologiaJoshuaKKyleNessuna valutazione finora
- Sistema NervosoDocumento15 pagineSistema NervosoBruce WiiNessuna valutazione finora
- Strumentazione BiomedicaDocumento15 pagineStrumentazione BiomedicaMarco AmatrudaNessuna valutazione finora
- Appunti Settimana 1 CBI ITADocumento10 pagineAppunti Settimana 1 CBI ITALuna SartoriNessuna valutazione finora
- MetodiDocumento7 pagineMetodiLuisaCurlacciNessuna valutazione finora
- Dossier GiulianiDocumento13 pagineDossier GiulianimodenafraNessuna valutazione finora
- OLIVIERO AUDISIO EstrattoDocumento8 pagineOLIVIERO AUDISIO EstrattoBiancamacarioNessuna valutazione finora
- Sinapsi e NeurotrasmettitoriDocumento45 pagineSinapsi e NeurotrasmettitoriNicola Laurora0% (1)
- Riassunto CitologiaDocumento38 pagineRiassunto CitologiaAlice QuadroNessuna valutazione finora
- Indice Kandel - Principi Di NeuroscienzeDocumento26 pagineIndice Kandel - Principi Di NeuroscienzeMarco CurottiNessuna valutazione finora
- Fondamenti Anatomo Fisiologici Dellattivita PsichicaDocumento29 pagineFondamenti Anatomo Fisiologici Dellattivita PsichicagigioNessuna valutazione finora
- FISIOLOGÍADocumento26 pagineFISIOLOGÍAAndrea De La FuenteNessuna valutazione finora
- Fisiologia SinapsiDocumento1 paginaFisiologia SinapsiFedericaRomanoNessuna valutazione finora
- Potenziale D'azioneDocumento30 paginePotenziale D'azioneRiccardo Mason TroiaNessuna valutazione finora
- Effetti Fisiologici Derivanti Dall'Assunzione Di MDMADocumento31 pagineEffetti Fisiologici Derivanti Dall'Assunzione Di MDMAhellenNessuna valutazione finora
- Il Potenziale D'azioneDocumento6 pagineIl Potenziale D'azioneLeandra EvolaNessuna valutazione finora
- Trasmissione Sinaptica e Integrazione NeuronaleDocumento11 pagineTrasmissione Sinaptica e Integrazione NeuronaleAnnamaria MaderaNessuna valutazione finora
- Anche Ad Altre ComponentiDocumento4 pagineAnche Ad Altre Componentifrancesco biancoNessuna valutazione finora
- FISIOLOGIADocumento30 pagineFISIOLOGIAlorenzo carliniNessuna valutazione finora
- 5 Potenziali ElettrotoniciDocumento8 pagine5 Potenziali ElettrotoniciClaudia OrlannoNessuna valutazione finora
- 4 Circuito Elettrico Equivalente Alla Membrana PlasmaticaDocumento6 pagine4 Circuito Elettrico Equivalente Alla Membrana PlasmaticaClaudia OrlannoNessuna valutazione finora
- 3 Equazione Di GoldmanDocumento2 pagine3 Equazione Di GoldmanClaudia OrlannoNessuna valutazione finora
- 2 Potenziale Di Equilibrio Di Gibbs-DonnanDocumento2 pagine2 Potenziale Di Equilibrio Di Gibbs-DonnanClaudia OrlannoNessuna valutazione finora
- Domande e Risposte Sistema NervosoDocumento4 pagineDomande e Risposte Sistema Nervosoedup220% (1)
- La Plasticita Cerebrale Aa 14.15Documento56 pagineLa Plasticita Cerebrale Aa 14.15Pia MisitiNessuna valutazione finora
- Lezione 10 SinapsiDocumento69 pagineLezione 10 SinapsiCopisteria Le PapierNessuna valutazione finora
- Esame FisiologiaDocumento17 pagineEsame FisiologiaelisabettaNessuna valutazione finora
- Neurologia 2020-2021Documento205 pagineNeurologia 2020-2021federicoNessuna valutazione finora
- Arrighi Superstar - Fondamenti Terzo Modulo Sistemato DefinitivoDocumento44 pagineArrighi Superstar - Fondamenti Terzo Modulo Sistemato Definitivoluigi cremascoliNessuna valutazione finora
- PsicostimolantiDocumento44 paginePsicostimolantiAlmy AlexandraNessuna valutazione finora
- Sistema Nervoso - ANATOMIA 2 PDFDocumento244 pagineSistema Nervoso - ANATOMIA 2 PDFvbnm,.Nessuna valutazione finora
- Integrazione Di Bcaa e SlaDocumento124 pagineIntegrazione Di Bcaa e SlamednasrallahNessuna valutazione finora
- Istologia Appunti - OdtDocumento20 pagineIstologia Appunti - Odtprova123100% (1)
- Psicofisiologia Di Apprendimento e MemoriaDocumento25 paginePsicofisiologia Di Apprendimento e Memoriatyrone_orozcoNessuna valutazione finora
- Psicologia GeneraleDocumento14 paginePsicologia GeneraleMATILDE MAISTONessuna valutazione finora
- NEUROLOGIADocumento23 pagineNEUROLOGIAracheleNessuna valutazione finora
- Neurobiologia Della MemoriaDocumento52 pagineNeurobiologia Della MemoriaanandhamideNessuna valutazione finora
- Anche Gli Alberi Hanno Un CervelloDocumento3 pagineAnche Gli Alberi Hanno Un CervelloLello GuarinoNessuna valutazione finora
- Tessuto Muscolare Striato ScheletricoDocumento26 pagineTessuto Muscolare Striato ScheletricoCinzia Modica100% (1)
- Neuroanatomia (PG)Documento82 pagineNeuroanatomia (PG)Sio88100% (2)
- Appunti Anatomia VeterinariaDocumento25 pagineAppunti Anatomia VeterinariasimoneNessuna valutazione finora
- Le Tattiche del Cambiamento - Manuale di Psicoterapia StrategicaDa EverandLe Tattiche del Cambiamento - Manuale di Psicoterapia StrategicaValutazione: 5 su 5 stelle5/5 (1)
- Piena Coscienza : Lezioni da leggere prima di dormireDa EverandPiena Coscienza : Lezioni da leggere prima di dormireNessuna valutazione finora
- Anatomia della Guarigione: I sette principi della Nuova Medicina IntegrataDa EverandAnatomia della Guarigione: I sette principi della Nuova Medicina IntegrataValutazione: 5 su 5 stelle5/5 (2)
- Il Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoDa EverandIl Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoNessuna valutazione finora
- Anatomia della Coscienza Quantica: La fisica dell'auto-guarigioneDa EverandAnatomia della Coscienza Quantica: La fisica dell'auto-guarigioneValutazione: 4.5 su 5 stelle4.5/5 (3)
- La medicina di santa Ildegarda: Per tutti i giorni e per tutta la famiglia.Da EverandLa medicina di santa Ildegarda: Per tutti i giorni e per tutta la famiglia.Valutazione: 5 su 5 stelle5/5 (2)
- Gonfiore Addominale: Guide e consigli per contrastare la pancia gonfiaDa EverandGonfiore Addominale: Guide e consigli per contrastare la pancia gonfiaNessuna valutazione finora
- Identità Rara. La lotta dei malati rari per il riconoscimento socialeDa EverandIdentità Rara. La lotta dei malati rari per il riconoscimento socialeNessuna valutazione finora
- Il miracolo della medicina rigenerativa: Come invertire il processo d’invecchiamento in modo naturaleDa EverandIl miracolo della medicina rigenerativa: Come invertire il processo d’invecchiamento in modo naturaleNessuna valutazione finora
- Prevenire le Malattie: Strategie Rivoluzionarie per un Domani più Sano: Potenziare il Tuo Percorso verso il Benessere attraverso la Scienza e l'InnovazioneDa EverandPrevenire le Malattie: Strategie Rivoluzionarie per un Domani più Sano: Potenziare il Tuo Percorso verso il Benessere attraverso la Scienza e l'InnovazioneNessuna valutazione finora
- Il Piano Dietetico del Dr. Nowzaradan: La Bilancia non Mente, la Gente Mente! L'unica Dieta da 1200 kcal del Dr NOW per Perdere Peso Velocemente. Piano Dietetico di 30 GiorniDa EverandIl Piano Dietetico del Dr. Nowzaradan: La Bilancia non Mente, la Gente Mente! L'unica Dieta da 1200 kcal del Dr NOW per Perdere Peso Velocemente. Piano Dietetico di 30 GiorniNessuna valutazione finora
- Ricomincia dalla pancia: Scopri il potere anti-età del microbioma e ritrova la tua salute partendo da dentroDa EverandRicomincia dalla pancia: Scopri il potere anti-età del microbioma e ritrova la tua salute partendo da dentroIsabella PolliNessuna valutazione finora
- La vera agopuntura cinese - Dottrina, Diagnosi, TerapiaDa EverandLa vera agopuntura cinese - Dottrina, Diagnosi, TerapiaNessuna valutazione finora
- Autodesk Inventor | Passo dopo Passo: Progettazione CAD e Simulazione FEM con Autodesk Inventor per PrincipiantiDa EverandAutodesk Inventor | Passo dopo Passo: Progettazione CAD e Simulazione FEM con Autodesk Inventor per PrincipiantiNessuna valutazione finora
- Nomofobia: Cos'è, chi ne soffre, perché esisteDa EverandNomofobia: Cos'è, chi ne soffre, perché esisteNessuna valutazione finora
- Biologia quantistica e Cronobiologia fasciale osteopaticaDa EverandBiologia quantistica e Cronobiologia fasciale osteopaticaNessuna valutazione finora