Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
Docsity Domande Triennale Fisiologia Umana
Caricato da
destinyema78Copyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
Docsity Domande Triennale Fisiologia Umana
Caricato da
destinyema78Copyright:
Formati disponibili
Domande Triennale Fisiologia
Umana
Fisiologia Umana
Università degli Studi G. d'Annunzio Chieti - Pescara
32 pag.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
FISIOLOGIA UMANA
1. COS’È IL POTENZIALE DI MEMBRANA A RIPOSO E DA COSA È DETERMINATO? DISTRIBUZIONE
DEGLI IONI E DELLE CARICHE FRA AMBIENTE ESTERNO ED INTERNO DELLA CELLULA.
Il potenziale di membrana è la differenza di potenziale elettrico tra il liquido intracellulare e il
liquido extracellulare. A riposo le cellule hanno un potenziale di membrana tale che l’interno
della cellula risulta più carico negativamente rispetto all’esterno dovuto dal fatto che
all’interno c’è una maggiore concentrazione di ioni negativi.
Il potenziale di membrana è determinato da forze elettrochimiche che spingono gli ioni di
potassio all’esterno e gli ioni sodio all’interno; a riposo la cellula è più permeabile al potassio e
quindi possiamo dire che questo potenziale è dovuto principalmente al potassio.
Una cellula ha un potenziale di membrana a riposa di -70 mV, considerando la cellula più
permeabile al potassio questo potenziale è più vicino al potenziale di equilibrio del potassio
(Ek) che è pari a -90 mV, che al potenziale del sodio (ENa) che è pari +60 mV.
Un altro motivo che determina il potenziale di membrana è l’azione della pompa sodio-
potassio che è la più importante proteina di trasporto che mantiene i gradienti di
concentrazione di sodio e potassio attraverso la membrana. 3 Na+ fuori dalla cellula e 2 K+ nella
cellula.
▪ L’elevata permeabilità della membrana al K+
▪ L’impermeabilità degli ioni proteici, che rimanendo all’interno determinano la
polarizzazione negativa all’interno della cellula
▪ Dalla scarsa permeabilità della membrana al Na+
▪ Dall’azione della pompa sodio-potassio che contrasta il modesto flusso entrante di Na+ e la
conseguente fuoriuscita di K+, mantenendo inalterati i rapporti di concentrazione degli ioni
a cavallo della membrana.
2. MI DEFINISCA E SPIEGHI IL CONCETTO DI DEPOLARIZZAZIONE E IPERPOLARIZZAZIONE.
Il potenziale di membrana a riposo si trova a un
valore stazionario di -70 mV. Quando la traccia
si muove verso l’alto (positivo), la differenza di
potenziale tra l’interno e l’esterno della cellula
diminuisce, si dice che la cellula è
depolarizzata. Se il potenziale diventa più
negativo, la differenza di potenziale aumenta e
si dice che la cellula si è iperpolarizzata.
Il ritorno del potenziale di membrana al valore
di riposo di -70 mV è detto ripolarizzazione.
In conclusione, per depolarizzazione si intende una diminuzione della negatività intracellulare
e per iperpolarizzazione invece si intende un aumento della negatività intracellulare.
3. IL NEURONE: CENTRO INTEGRATIVO DI SEGNALI NERVOSI
Trasportano velocemente i segnali elettrici e, in alcuni casi, su grandi distanze. I neuroni
presentano un corpo o soma da cui escono lunghi processi chiamati dendriti se ricevono
segnali in arrivo o assoni se trasportano informazioni in uscita.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Il corpo è il centro di controllo del neurone ed ha una confermazione simile a tutte le cellule
con un nucleo e tutti gli organuli necessari per l’attività cellulare.
I dendriti sono lunghi processi ramificati che ricevono informazioni in entrata da cellule vicine.
La loro funzione principale nel SNP è portare l’informazione in direzione centripeta (periferia
→ centro)
Gli assoni invece trasportano i segnali in uscita fino agli organi bersaglio in direzione centrifuga
(centro → periferia)
Le cellule nervose sono descritte come tessuti eccitabili per la loro capacità di produrre e
propagare rapidamente segnali elettrici in risposta ad uno stimolo.
4. COSA È – E QUALI SONO LE CARATTERISTICHE – DI UN POTENZIALE GRADUATO?
I potenziali graduati sono detti così perché la sua intensità è direttamente proporzionale
alla forza dell’evento scatenante per questo sono segnali ad intensità variabile che si
propagano per brevi distanze e man mano che si allontano dal punto in cui sono stati
generati perdono di intensità.
Quando più potenziali graduati si combinano in maniera simultanea è detta sommazione
spaziale ed è riferito al fatto che essi hanno origine in punti diversi del neurone. La
sommazione di potenziali graduati non richiede input.
Quando due potenziali graduati sottosoglia provenienti dallo stesso neurone arrivano alla
zona trigger abbastanza vicini nel tempo si sovrappongono e si ha una sommazione
temporale.
Nei neuroni i potenziali graduati hanno luogo quando i segnali chimici proveniente da altri
neuroni aprono i canali ionici regolati chimicamente permettendo agli ioni di entrare o
uscire dal neurone; possono verificarsi anche quando un canale aperto si chiude,
diminuendo il movimento di ioni attraverso la membrana.
Quando ioni Na+ si spostano all’interno della cellula introducono carica elettrica positiva
che si diffonde come un’onda di depolarizzazione attraverso il citoplasma ed essa è
associata ad un flusso locale di carica.
L’intensità della depolarizzazione iniziale in un potenziale graduato è determinata da
quanta carica penetra nella cellula; quanto più grande è l’ampiezza iniziale tanto più
lontano si diffonderà lungo il neurone prima di esaurirsi.
Il potenziale graduato perde di intensità:
a. La dispersione di corrente nel liquido extracellulare perché ci sono canali aperti che
permettono alla carica positiva di disperdersi.
b. La resistenza del citoplasma che si oppone al flusso di corrente.
I potenziali graduati abbastanza intensi raggiungono la zona trigger (centro di integrazione
del neurone), che contiene un’alta concentrazione di canali Na+ voltaggio-dipendenti, e se
depolarizzano la membrano fino al livello soglia (-55 mV) i canali si aprono e innescano un
potenziale d’azione.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
5. COSA SI INTENDE PER POTENZIALE GRADUATO SOTTOSOGLIA O SOVRASOGLIA?
Se un potenziale graduato in depolarizzazione è abbastanza intenso quando raggiunge la
zona trigger allora riuscirà ad innescare un potenziale d’azione è detto sovrasoglia. Invece
un potenziale graduato sottosoglia non raggiunge la zona trigger di un neurone perché la
sua intensità è inferiore al valore soglia (-55 mV) e quindi non riesce ad innescare un
potenziale d’azione.
6. POTENZIALE D’AZIONE NEL NEURONE: GRAFICO E SPIEGAZIONE DEI MOVIMENTI IONICI.
Un potenziale d’azione non perde di intensità
quindi all’estremità di un assone avrà la stessa
intensità di quando parte dalla zona trigger per
questo viene definito come un fenomeno “tutto
o nulla” perché si presenta con una
depolarizzazione massima oppure non si
presenta affatto.
Esso può essere suddiviso in tre fasi:
a. Fase ascendente dovuta ad un improvviso aumento della permeabilità della cellula al
sodio. Quando un potenziale graduato arriva alla zona trigger depolarizza la membrana
fino al livello soglia e i canali per il sodio voltaggio-dipendenti si aprono e rendono essa
molto più permeabile. Dato che l’interno della cellula è più negativo dell’esterno attrae
ioni sodio con carica positiva e l’aggiunta di tale carica fa venir meno il gradiente
elettrico; tuttavia il gradiente di concentrazione del sodio rimane quindi questi ioni
continuano ad entrare nella cellula cercando di arrivare al suo potenziale di equilibrio
(+60 mV). La fase di picco viene raggiunta però quando il potenziale di azione arriva ad
un valore di +30 mV, poiché i canali del sodio nell’assone si chiudono e quelli del
potassio si aprono.
b. Fase discendente corrisponde ad un aumento della permeabilità del potassio. I canali
per il potassio voltaggio-dipendenti si aprono in risposta alla depolarizzazione, però
essendo più lenti il picco arriva più tardi. Quando il potenziale di membrana è positivo
viene meno il gradiente elettrico e di concentrazione favorendo la fuoriuscita del
potassio dalla cellula, questo fa si che il potenziale diventa negativo creando la fase
discendente del potenziale d’azione.
c. Fase di iperpolarizzazione postuma. Quando il potenziale di membrana discendente
raggiunge il -70 mV i canali per il potassio non si sono ancora chiusi, quindi la
membrana si iperpolarizza avvinandosi al valore di equilibrio del potassio (-90 mV).
Infine, i canali lenti si chiudono, la fuoriuscita del potassio cessa e grazie alla ritenzione
e la penetrazione del sodio il potenziale di membrana torna al valore di riposo di -70
mV.
7. I PERIODI REFRATTARI: IL RUOLO DEI CANALI VOLTAGGIO DIPENDENTI DAL SODIO.
Durante la fase di depolarizzazione la cellula eccitabile non può essere nuovamente
attivata per circa 1-2 ms quindi si dice che si trova in periodo refrattario (resistente)
assoluto. Questo è dato dall’impossibilità di far entrare ulteriore sodio per via
dell’inattivazione dei suoi canali.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Esistono due tipi di periodi refrattari:
a. Periodo refrattario assoluto che inizia a -55 mV e finisce quando inizia
l’iperpolarizzazione postuma
b. Periodo refrattario relativo che può generare un secondo potenziale d’azione solo
applicando stimoli con intensità superiore a quella che ha generato il potenziale
precedente. Esso è tanto più lungo quanto più è presente l’iperpolarizzazione postuma.
I canali voltaggio-dipendenti reagiscono ai cambiamenti nel potenziale di membrana nella
cellula ed hanno un ruolo fondamentale nella generazione della conduzione dei segnali
elettrici. Quelli del sodio hanno due cancelli: uno di attivazione e uno di inattivazione che
regolano il passaggio di ioni. Quando il potenziale di membrana si trova a riposo il cancello
attivazione del canale per il sodio è chiuso mentre quello di inattivazione è aperto. Se la
membrana cellulare vicino al canale si depolarizza il cancello di attivazione si apre perché
sia il cancello di attivazione che di inattivazione si muovono in risposta alla
depolarizzazione l’unica differenza è il ritardo del secondo di circa 0.5 ms rispetto al primo;
durante questo intervallo il canale è aperto permettendo l’ingresso di ioni sodio tale da
generare la fase ascendente del potenziale d’azione. Raggiunto il picco del potenziale
d’azione i cancelli si chiudono, il sodio smette di entrare, il potassio esce, la cellula si
ripolarizza e si riprepara per la depolarizazione successiva.
8. CONDUZIONE SALTATORIA NELL’ASSONE.
Gli assoni mielinizzati limitano la quantità di membrana a contatto conil fluido
extracellulare lasciando porzioni di membrana scoperta, i nodi di Ranvier, che si alternano
con segmenti lunghi avvolti in strati multipli di membrana (guaina mielinica). Quest’ultima
crea una parete molto resistente che impedisce al flusso di ioni di uscire dal citoplasma. Il
processo di conduzione si verifica solo a livello dei nodi e ognuno di essi presenta un’alta
concentrazione di canali sodio voltaggio-dipendenti, che si aprono con la depolarizzazione
lasciando entrare il sodio nell’assone; rinforzano e mantengono costante l’ampiezza del
potenziale d’azione mentre questo passa da un nodo all’altro. Questo andamento si dice
saltatorio proprio perché sembra “saltare”.
9. LA GIUNZIONENEUROMUSCOLARE: STRUTTURA E FUNZIONE.
Una giunzione neuromuscolare è la sinapsi di un motoneurone somatico su una fibra
muscolare, composta da:
a. Terminale assonico del neurone presinaptico motorio riempito di vescicole sinaptiche e
mitocondri
b. Fessura sinaptica
c. Membrana postsinaptica della fibra muscolare scheletrica
Inoltre, comprende estensioni delle cellule di Schwann che formano uno strato sottile di
rivestimento per le estremità dei terminali assonici.
Sul lato postsinaptico la zona di membrana localizzata di fronte al terminale assonico viene
modificata e va a formare una placca motrice che contiene un’alta concentrazione di
recettori nicotinici per l’ACh. La fessura sinaptica è occupata da una matrice fibrosa, le cui
fibre mantengono il terminale assonico e la placca motrice correttamente allineati.
l’ACh, legandosi ai recettori nicotinici presenti nella placca motrice, apre i canali per il Na +,
l’ingresso netto di sodio nella fibra muscolare depolarizza la fibra stessa.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
10. ORGANIZZAZIONE GENERALE DEL SISTEMA NERVOSO CENTRALE E QUALI SONO LE SUE
FUNZIONI.
Il SNC è costituito dall’encefalo e dal midollo spinale. Ai primi stadi embrionali le cellule da
cui si originerà il SNC si trovano in una regione appiattita chiamata placca neurale, che
mostra due pieghe laterali dette creste neurali, con lo sviluppo i bordi della placca si
fondono tra loro creando un tubo neurale. Dalla sesta settimana il SNC è diviso in 7
porzioni principali: sei di queste parti sono il telencefalo, il diencefalo, il mesencefalo, il
cervelletto, il ponte e il bulbo o midollo allungato, la settimana è il midollo spinale. In
seguito, il tubo neurale si inizia ad allargare formando i ventricoli cerebrali.
Il SNC è costituito da neuroni e cellule gliali di sostegno e da due sostanze una bianca ed
una grigia. La materia grigia non contiene mielina ed è costituita dai corpi cellulari delle
cellule nervose, dai dendriti e dalle parti terminali degli assoni. La materia bianca invece è
principalmente costituita da assoni mielinizzati (che scaturisce il colore bianco) e contiene
pochi corpi cellulari. Tra il tessuto osseo e il SNC ci sono 3 membrane chiamate meningi
utili a proteggere il tessuto osseo dalle contusioni; queste membrane sono, partendo
dall’esterno verso l’interno, dura madre – aracnoide e pia madre.
Il SNC riceve e analizza le informazioni in arrivo dall’ambiente interno ed esterno
dell’organismo per poi elaborarle e rilasciare risposte appropriate.
11. RUOLO DI MENINGI E LIQUIDO CEREBROSPINALE NELLA PROTEZIONE DEL SISTEMA NERVOSO
Le meningi sono 3:
▪ DURA MADRE è la più spessa e contiene i vasi venosi che drenano il sangue grazie a delle
cavità chiamate seni
▪ ARACNOIDE connessa con la membrana più interna e contiene uno spazio subaracnoideo
▪ PIA MADRE più interna e sottile, essa aderisce alla superficie dell’encefalo e del midollo
spinale, contiene vasi arteriosi che irrorano l’encefalo.
Il LIQUIDO CEREBROSPINALE (LCS) è una soluzione salina secreta dai plessi corioidei ed ha la
funzione di protezione chimica e fisica e fornisce una sorta di imbottitura protettiva dagli urti
(assimilabile ad un ammortizzatore idraulico).
12. DIFFERENZE ANATOMICHE E FUNZIONALI FRA IL SISTEMA NERVOSO SIMPATICO E
PARASIMPATICO.
Il sistema nervoso autonomo regola tutte le attività dei nostri organi che non sono
comandati direttamente dalla nostra volontà ed è diviso in 2 componenti: il sistema
simpatico che interviene nelle situazioni di emergenza, e il sistema parasimpatico che
agisce nei momenti di relax. Le principali differenze sono:
a. le origini delle vie nel SNC
b. la localizzazione dei gangli
la maggior parte delle vie simpatiche origina a livello della regione toracica e lombare del
midollo spinale. I gangli simpatici sono situati in 2 catene che decorrono parallelamente ai
lati della colonna vertebrale. Esse di solito hanno assoni pregangliari brevi e assoni
postgangliari lunghi. Invece, molte vie parasimpatiche originano nel tronco encefalico e
lasciano il SNC tramite alcuni nervi cranici. Altre vie originano nella regione sacrale e
controllano gli organi pelvici. L’innervazione parasimpatica riguarda primariamente la testa
il collo e gli organi interni.
Dal punto di vista funzionale i sistemi simpatico e parasimpatico vengono distinti in base ai
rispettivi neurotrasmettitori e recettori:
- i neuroni autonomi post gangliari sia simpatici che parasimpatici secernono acetilcolina su
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
recettori colinergici nicotinici localizzati sui neuroni post gangliari;
- la maggior part dei neuroni simpatici rilascia noradrenalina sui recettori adrenergici delle
cellule bersaglio;
- i neuroni parasimpatici rilasciano ACh sui recettori colinergici delle cellule bersaglio.
I principali neurotrasmettitori dell’SNA sono acetilcolina e noradrenalina facilmente
sintetizzate dagli enzimi citoplasmatici.
In breve:
1) le vie simpatiche e parasimpatiche sono entrambe costituite da due neuroni in serie
(pregangliari e postgangliari).
2) tutti i neuroni autonomi pregangliari secernono acetilcolina → agisce sui recettori
nicotinici. I neuroni simpatici postgangliari secernono noradrenalina → agisce sui recettori
adrenergici. I neuroni parasimpatici postgangliari secernono acetilcolina → agisce sui
recettori muscarinici.
3) vie simpatiche originano dalle regioni toraciche e lombari del midollo spinale e i gangli si
trovano vicino al midollo spinale. I neuroni parasimpatici lasciano il SNC a livello del tronco
encefalico e della regione sacrale del midollo spinale e i suoi gangli sono in prossimità dei
tessuti bersaglio.
4) il sistema simpatico controllo funzioni utili soprattutto nelle situazioni di stress e di
emergenza. Il sistema parasimpatico è dominante durante il riposo e la digestione.
Quando è in azione il sistema simpatico l’organismo consuma molta energia, infatti il
fegato converte il glicogeno di riserva in glucosio per fornire energia. Invece, quando è in
azione il sistema parasimpatico l’organismo si rilassa e accumula l’energia sotto forma di
sostanze di riserva.
13. SIMILARIITÀ E DIFFERENZE TRA GIUNZIONI NEURO-MUSCOLARE E NEURO-EFFETRICE
▪ Neuroeffettrice: sinapsi tra il neurone autonomico e le sue cellule bersaglio
▪ Neuromuscolare: sinapsi di un motoneurone somatico su una fibra muscolare.
14. IL MECCANISCO DI ACCOPPIAMENTO ECCITAZIONE CONTRAZIONE (EC) NEL MUSCOLO
SCHELETRICO: QUALI SISTEMI DI MEMBRANE E QUALI PROTEINE SONO IMPLICATE.
È l’insieme dei processi grazie ai quali l’eccitazione elettrica della membrana avvia la
contrazione di strutture all’interno della cellula muscolare provocando l’aumento della
concentrazione di calcio intracellulare che a sua volta avvia il ciclo di contrazione-
rilasciamento.
Durante la contrazione agiscono due proteine l’actina e la miosina.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
L’accoppiamento eccitazione-contrazione inizia con il rilascio dell’acetilcolina da parte del
motoneurone somatico a livello della giunzione neuromuscolare, l’ACh si lega ai canali
chemio-dipendenti a livello della placca motrice facendoli aprire consentendo sia al Na+
che al K+ di attraversare il sarcolemma. Queste cariche positive depolarizzano la membrana
dando origine ad un POTENZIALE DI PLACCA (PP). Il potenziale d’azione si propaga sulla
superfice della cellula e nei tubuli T grazie all’apertura dei canali del Na+ voltaggio-
dipendenti, avviene similmente a quella nelle fibre nervose ma nella fibra muscolare è più
lenta.
15. QUANTI E QUALI TIPI DI MUSCOLO SONO PRESENTI NEL NOSTRO ORGANISMO? SIMILIRATÀ E
DIFFERENZE FRA DI ESSI.
Caratteristiche Muscolo Muscolo Muscolo liscio
Scheletrico cardiaco
Tipo di cellule Polinucleate Mononucleate Mononucleate
Componenti Endomisio
connettivali Epimisio attaccato allo
Perimisio scheletro Solo Endomisio
Endomisio filamentoso del
cuore
Presenza di Si, ma le No, ma i
miofibrille costituite Si miofibrille filamenti di
da sarcomeri hanno spessore actina e miosina
irregolare sono presenti
Presenza di tubuli T Si, 2 in ciascun Si a livello delle No, ma caveole
sarcomero a livello linee Z con lungo il
delle giunzioni A e I diametro sarcolemma
maggiore
Reticolo Meno del Come nel
sarcoplasmatico (RS) muscolo cardiaco, e il RS
Si scheletrico, è a contatto con
poche cisterne il sarcolemma
terminali
Presenza di giunzioni No Si a livello dei Si nel muscolo
comunicanti dischi intercalari unitario
Cellule che No, nel muscolo
innervano giunzioni Si No unitario
neuromuscolari Si, nel muscolo
multiunitario
Fonte di Calcio per la RS RS e fluido RS e fluido
contrazione extracellulare extracellulare
16. IL MUSCOLO SCHELETRICO: RELAZIONE TRA FIBRA MUSCOLARE, MIOFIBRILLA E SARCOMERO.
Il muscolo scheletrico è l’insieme di fibre muscolari (lunghe cellule polinucleate). Ognuna di
esse contiene delle cellule satelliti che si trovano appena all’esterno della loro membrana,
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
si attivano e si differenziano quando è necessario per la crescita e la riparazione del
muscolo. Le fibre sono avvolte da tessuto connettivo che raggruppa quelle adiacenti
formando dei fascicoli.
La membrana della fibra muscolare è detta sarcolemma e il citoplasma è detto
sarcoplasma. La principale struttura intracellulare è rappresentata dalle miofibrille
(struttura contrattile della fibra muscolare). Le fibre anche un esteso reticolo
sarcoplasmatico che avvolge ciascuna miofibrilla ed è costituito da tubuli longitudinali
chiamati cisterne terminali associate ad una rete ramificata di tubuli trasversi, Tubuli T.
Ogni miofibrilla è costituita da diversi tipi di proteina:
• Proteine contrattili, miosina e actina
• Proteine regolatrici, tropomiosina e troponina
• Proteine strutturali, titina e nebulina.
Le miofibrille sono costituite da sarcomeri posti in serie.
17. PROTEINE DEL SARCOMERO: QUANTI E QUALI? ORGANIZZAZIONE E FUNZIONI.
Le proteine che troviamo all’interno del sarcomero sono:
• Miosina: ha la capacità di generare movimento e costituisce il filamento spesso del
sarcomero
• Actina: costituisce il filamento sottile
o I filamenti spessi e sottili rappresentano le parti contrattili del sarcomero,
l’allineamento corretto di questi è assicurato da delle proteine strutturali:
• Titina: proteina elastica che
si estende dalla linea Z al
centro del filamento di
miosina ed ha la funzione di
(1) stabilizzare la posizione
dei filamenti contrattili e (2)
consentire il ritorno del
muscolo alla sua lunghezza
iniziale dopo lo stiramento.
• Nebulina: proteina
anelastica che si estende
lungo i filamenti sottili e si
attacca alle linee Z e
favorisce l’allineamento dei
filamenti di actina.
• Ogni sarcomero è:
• Delimitato da due linee Z successive
costituite da proteine che servono da
attacco per i filamenti sottili;
Alle estremità ci sono delle bande più
chiare chiamate bande I composte da
filamenti sottili e sono attraversate a
metà dalla linea Z e ogni metà delle
bande I appartiene a due sarcomeri
diversi.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Bande A più scure e coprono tutta la lunghezza del filamento spesso, alle estremità i
filamenti spessi e sottili si sovrappongono.
Zona H regione centrale dove si trovano solo filamenti spessi di miosina.
Linea M divide a metà la banda A ed è costituita da proteine accessorie a cui si attaccano i
filamenti spessi.
18. LA MIOSINA: ENZIMA, MOTORE MOLECOLARE E PROTEINA STRUTTURALE.
La miosina è considerato un motore molecolare perché possiede la capacità di generare
movimento. Ciascuna molecola è costituita da catene proteiche che si intrecciano a
formare una lunga coda con due teste globulari che hanno una regione elastica che fa da
cardine nel punto di unione con la coda che è rigida. Ogni testa ha due catene proteiche:
una catena pesante ed una leggera, la catena pesante rappresenta il dominio motore che
lega l’ATP e usa l’energia del legame fosfato per generare movimento. Oltre al sito per
l’ATP la catena pesante ha un altro sito per l’actina.
Il dominio motore agisce come un enzima ed è considerata una miosina ATPasi in grado di
idrolizzare ATP in ADP e fosfato inorganico.
La flessione delle teste di miosina legate all’actina produce la forza che fa scorrere i
filamenti di actina durante la contrazione.
“questo scorrimento spiega come un muscolo si possa contrarre e generare forza senza
movimento”
Le teste di miosina si legano alle molecole di actina, un segnale mediato dal calcio avvia il
“power stroke”, durante il quale il ponte di miosina tira il filamento di actina verso il centro
del sarcomero. Alla fine ciascuna testa di miosina si stacca dall’actina, si ridistende e si
riattacca ad un’altra molecola per iniziare un nuovo ciclo di contrazione. Le teste non si
rilasciano tutte allo stesso tempo, altrimenti la fibra tornerebbe alla sua lunghezza iniziale.
Gli eventi molecolari di un ciclo di contrazione iniziano dallo stato di rigor, uno stato in cui
le teste testa della miosina sono strettamente legate alle molecole di G-actina.
(1) ATP si lega e la miosina si distacca
Una molecola di ATP si lega al sito della miosina, il legame ATP-miosina diminuisce
l’affinità di legame dell’actina per la miosina che quindi si distacca.
(2) L’idrolisi dell’ATP fornisce l’energia per l’estensione della testa di miosina e il suo
riattacco al sito dell’actina
L’energia liberata dall’ATP ruota la testa della miosina fino a formare un angolo di 90° e
si lega debolmente a un nuovo sito di actina.
(3) Il “power stroke”
Inizia dopo che il calcio si è legato alla troponina, questo legame fa spostare la
tropomiosina che libera completamente i siti di legame.
(4) La miosina rilascia ADP al termine del colpo di forza
La miosina è una proteina strutturale perché fa parte della struttura del sarcomero e ne
costituisce il filamento spesso.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
19. LE PROTEINE REGOLATRICI TROPONINA E TROPOMIOSINA: LA LORO FUNZIONE.
Il segnale che avvia o arresta il processo di contrazione è l’aumento o la diminuzione della
concentrazione di calcio intracellulare. La troponina è la proteina legante il calcio e
controlla la posizione della tropomiosina. A riposo la tropomiosina si avvolge attorno
all’actina coprendo parzialmente i siti di legame per la miosina, questa posizione è detta
“off” o di blocco. Prima che avvenga la contrazione si passa nella posizione “on” in cui si
liberano i siti di legame e questo cambiamento è controllato dalla troponina.
In presenza di calcio la troponina lega reversibilmente il calcio, questo legame sposta la
molecola di tropomiosina liberando i siti attivi per l’actina. In posizione “on” le teste di
miosina creano legami forti e avviano il “power stroke”.
La contrazione termina quando inizia a diminuire il Ca2+ intracellulare perché in assenza di
calcio la troponina permette alla tropomiosina di tornare nella posizione off coprendo
nuovamente i siti dell’actina per la miosina.
20. COS’È UN’UNITÀ MOTORIA? MI PARLI DI ATTIVITÀ MOTORIA E RECLUTAMENTO.
Un’unità motoria è l’unità fondamentale per la
contrazione del muscolo scheletrico, è costituita
da un motoneurone somatico e delle fibre
muscolari che esso innerva. Ogni fibra è innervata
da un solo motoneurone. Il numero di fibre che
costituisce un’unità motoria è molto variabile.
L’unità motoria è costituita da fibre tutte dello
stesso tipo, per questo si parla di unità motorie
lente o rapide.
Ogni unità motoria si contrae in maniera tutto o
nulla. Il muscolo riesce a generare contrazioni di
forza e durata diversa perché è costituito da molte
unità motorie (1) reclutando tipi diversi di unità motorie e (2) cambiando il numero di unità
motorie attivate in un determinato momento.
La forza di contrazione può essere aumentata reclutando nuove unità motorie questo
reclutamento è sotto il controllo del sistema nervoso che procede in maniera stereotipata.
Uno stimolo poco intenso attiva soltanto i motoneuroni che hanno la soglia di eccitazione
bassa che controllano le fibre lente, all’aumentare del segnale eccitatorio vengono attivati
altri motoneuroni a soglia più alta che fanno capo a unità motorie costituite da fibre
ossidative resistenti alla fatica. Quando aumenta ancora i motoneuroni a soglia più alta,
che stimolano unità motorie costituite da fibre più rapide e glicolitiche e la forza sviluppata
dal muscolo è vicina al suo valore massimo.
Una contrazione forte ha bisogno di una serie di potenziali d’azione dal SNC al muscolo.
Aumentando la frequenza si ha la sommazione delle contrazioni, ma se la fibra si affatica
facilmente questa stimolazione ne accelera l’affaticamento e quindi la riduzione di forza.
Un modo per evitare la fatica in una contrazione sostenuta è il reclutamento asincrono, il
SN modula il reclutamento dei motoneuroni così che diverse unità motorie a turno
mantengono la tensione muscolare. L’alternanza nel reclutamento delle unità motorie
permette ad alcune di esse di riposarsi tra le contrazioni, prevenendo l’affaticamento.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
21. CLASSIFICAZIONE DELLE FIBRE MUSCOLARI IN BASE AL METABOLISMO ED ALLA VELOCITÀ DI
CONTRAZIONE: RELAZIONE CON LE CAPACITÀ ATLETICHE DI UN INDIVIDUO.
Le fibre muscolari possono essere classificate sulla base delle loro velocità di contrazione e
sulla resistenza alla fatica.
• Fibre lente (tipo 1)
• Fibre rapide ossidativo-glicolitiche (tipo 2A)
• Fibre glicolitiche a contrazione rapida (tipo 2X nell’uomo, 2B nel topo)
Le fibre muscolari a contrazione rapida (tipo 2) sviluppano tensione 2-3 volte più
velocemente delle fibre a contrazione lenta (tipo 1). La velocità con cui una fibra muscolare
si contrae è determinata dall’isoforma della miosina ATPasi del filamento spesso. Le fibre
veloci scindono l’ATP più velocemente e possono quindi completare il ciclo di contrazione
più rapidamente.
La contrazione delle fibre rapide dura solo 7.5 ms invece quella delle fibre lente può durare
fino a 10 volte più a lungo, per questo le fibre rapide vengono usate raramente mentre
quelle lente costantemente per il mantenimento della postura e per la locomozione.
Le fibre glicolitiche (veloci di tipo 2B) utilizzano principalmente la glicolisi anaerobica per
rifornirsi di ATP e qui avremo uno sviluppo notevole della fatica, quindi si affaticano più
facilmente rispetto a quelle ossidative (tipo 1/ tipo 2A). Le fibre ossidative utilizzano
soprattutto la fosforilazione ossidativa per produrre ATP, queste fibre includono sia le fibre
lente (rosse) che quelle ossidative-glicolitiche.
Le fibre lente vengo anche chiamate rosse perché hanno un elevato contenuto di
mioglobina, un pigmento rosso con alta affinità per l’ossigeno e facilita la diffusione
all’interno delle fibre muscolari. L’elevato contenuto di mioglobina unito al piccolo diametro
di queste fibre rende la diffusione di ossigeno molto rapido e questo comporta il
mantenimento di un buon rifornimento di ossigeno che permette alle fibre ossidative di
produrre ATP tramite la fosforilazione ossidativa.
Le fibre glicolitiche sono dette bianche perché hanno un contenuto basso di mioglobina ed
hanno anche un diametro più grande rispetto a quelle ossidative. Questa combinazione
implica che le fibre glicolitiche vanno più incontro a scarsa disponibilità di ossigeno, quindi
per produrre ATP utilizzano principalmente la glicolisi anaerobica e si affaticano più
rapidamente.
Le fibre ossidative-glicolitiche hanno proprietà sia ossidative che glicolitiche, utilizzano una
combinazione di metabolismo ossidativo glicolitico ed hanno un diametro intermedio.
“Le fibre lente sono quelle che atrofizzano di più, perciò senza allenamento perdono prima la
massa”.
22. RELAZIONE TRA EVENTI ELETTRICI ED EVENTI MECCANICI NELLA FIBRA MUSCOLARE: COSA È
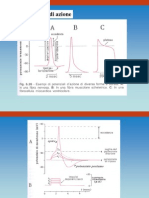
UNA SCOSSA SEMPLICE E MI PARLI DELLA SOMMAZIONE DEL TETANO.
Un potenziale d’azione proveniente dal SNC depolarizza
la membrana della fibra muscolare, questo evento
elettrico permette la contrazione del muscolo stesso. La
contrazione del muscolo costituisce l’evento meccanico.
La scossa semplice (single twich, a) è la risposta del
muscolo ad un singolo impulso, si può dire che è una
misurazione della forza.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
La prima fase della scossa semplice è il periodo di latenza
(arrivo del potenziale/generazione di forza). Segue la fase
di salita, cioè la contrazione, seguita da una fase di discesa
che rappresenta il rilassamento del muscolo. La fase di
salita e discesa sono controllati dalla concentrazione di
calcio. La tensione generata da una scossa semplice non
rappresenta la massima tensione che una fibra può
generare. Infatti, la forza può essere aumentata
incrementando la frequenza dei potenziali d’azione che la
stimolano.
Se gli stimoli sono ravvicinati, la fibra muscolare non si
rilascia del tutto e sviluppa una tensione maggiore per
l’effetto della sommazione delle risposte, questo processo si
chiama appunto sommazione (b).
Se i potenziali d’azione sono molto ravvicinati, ad alta
frequenza di stimolazione, l’entità del rilasciamento si riduce
e la fibra muscolare arriva ad uno stato di contrazione
chiamato tetano. I tipi di tetano sono 2: incompleto (c), in
cui la frequenza di stimolazione non è massimale e la fibra si
rilascia leggermente tra uno stimolo e l’altro; completo (d),
in cui la frequenza è sufficientemente alta da non dare tempo alla fibra di rilasciarsi e si
sviluppa la massima tensione possibile.
23. DEFINIZIONE E SIGNIFICATO DI CONTRAZIONE ISOMETRICA ED ISOTONICA NEL MUSCOLO
SCHELETRICO.
I muscoli generano forza per ottenere movimento, ma possono anche generare forza senza
produrre movimento.
La contrazione isotonica è una contrazione in cui c’è generazione di forza (costante) e
movimento di un carico. Durante questo tipo di contrazione è possibile ottenere un altro tipo
di evento, contrazione eccentrica. Quest’ultima è ritenuta una delle principali cause di danno
cellulare e di ritardo nel recupero dopo un esercizio fisico.
La contrazione isometrica è un tipo di contrazione che genera forza, ma non genera
movimento. Una contrazione isometrica può generare forza anche senza cambiare la
lunghezza del muscolo per la presenza di elementi elastici al suo interno. Nella contrazione
isometrica quando i sarcomeri si accorciano gli elementi elastici si allungano e questo
stiramento permette alla fibra di mantenere una lunghezza relativamente costante. Quando
gli elementi elastici sono stirati e la forza generata dai sarcomeri è uguale al peso, il muscolo
si contrae in maniera isotonica e solleva il carico.
In ogni contrazione c’è sempre una fase isometrica che può essere seguita da una fase
isotonica.
24. CURVA DI REGOLAZIONE TENIONE – LUNGHEZZA DEL SARCOMERO.
La tensione sviluppata da una fibra muscolare durante una contrazione dipende dalla
lunghezza di partenza dei sarcomeri all’inizio della contrazione. Un sarcomero sviluppa la sua
forza massima se si trova ad una lunghezza di lavoro ottimale.
Alla normale lunghezza a riposo dei muscoli, i sarcomeri si trovano vicino alla loro lunghezza
ottimale.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
La lunghezza del sarcomero è in stretta relazione con il grado di sovrapposizione tra filamenti
spessi e sottili.
La tensione che la fibra può generare è direttamente proporzionale al numero dei legami tra
le teste di miosina e i siti di actina. Se la contrazione inizia a lunghezze troppo grandi, i
filamenti spessi e sottili sono sovrapposti soltanto in piccola parte e formano ponti
trasversali.
Alla lunghezza ottimale si può formare il massimo numero di ponti trasversali tra filamenti
spessi e sottili e la fibra riesce a generare la massima forza. Se il sarcomero all’inizio
dell’azione è più corto i filamenti
sono troppo sovrapposti.
Se il sarcomero è così corto che i
filamenti arrivano a livello della
linea Z, le testa di miosina non
trovano nuovi siti di legame e la
tensione si riduce rapidamente.
Lo sviluppo della tensione
durante la scossa semplice è una
proprietà che dipende dalla
lunghezza del sarcomero.
25. IL MUSCOLO LISCIO: STRUTTURA E FUNZIONE.
Il muscolo liscio è il muscolo principale degli organi interni. È costituito da cellule
mononucleate e non sono striate, ha gli stessi elementi contrattili (proteine) del muscolo
scheletrico, però differenza di quest’ultimo i suddetti elementi non sono altamente
organizzati; infatti, nel muscolo liscio non sono presenti i sarcomeri.
Ci sono molti modi per classificare i diversi tipi di muscolo liscio:
▪ A seconda della sede in:
o Vascolare o Oculare
o Gastrointestinale o Respiratorio
o Urinario o Riproduttore
▪ A seconda del modello di contrazione, in funzione del fatto che alterni stati di rilasciamento
e di contrazione o se presenta uno stato di continua contrazione.
o Muscoli lisci fasici, sono i muscoli sottoposti a cicli di contrazione e rilasciamento
periodici. Es. Esofago si contrae solo quando il cibo lo attraversa.
o Muscoli lisci tonici, sono muscoli che sono continuamente contratti, perché devono
sempre mantenere un certo livello di tono muscolare.
▪ A seconda di come avviene la comunicazione con le cellule vicine
o Muscolo liscio unitario, muscoli in cui le cellule sono elettricamente collegate
attraverso gap junction e si contraggono come unità coordinate.
o Muscolo liscio multi – unitario, muscoli in cui le cellule non sono accoppiate
elettricamente e ciascuna cellula muscolare ha funzionamento autonomo. Per
questo ogni fibra può anche essere stimolata indipendentemente.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Nel muscolo liscio la forza è generata dall’interazione dei ponti trasversali di actina-miosina
e la contrazione è innescata da un aumento delle concentrazioni citosoliche di calcio libero.
Per altri aspetti, il funzionamento del muscolo liscio è più complesso:
▪ Devono operare dentro una gamma di lunghezze
▪ All’interno di un organo gli strati possono avere orientamenti diversi
▪ La contrazione e il rilasciamento avvengono in maniera più lenta
▪ Utilizzano meno energia per generare e mantenere una determinata quantità di
forza
▪ Hanno cellule piccole, fusiformi e mononucleate
▪ Le fibre contrattili non sono disposte in sarcomeri
▪ La contrazione può essere innescata da segnali elettrici o chimici, o da entrambi
▪ È controllato dal sistema nervoso autonomo
▪ Il calcio per la contrazione proviene dal liquido extracellulare
Nel muscolo liscio il segnale di inizio della contrazione è l’aumento del calcio citosolico, il
calcio si lega alla calmodulina, questo legame rappresenta il primo step e susseguono altri
eventi che terminano con la fosforilazione della catena leggera della miosina e questo
aumenta l’attività ATPasica e porta alla contrazione.
26. IL RUOLO DELLA CALMODULINA NELL’ATTIVAZIONE DELLA CONTRAZIONE DEL MUSCOLO
LISCIO.
La calmodulina è una proteina che lega il calcio presente nel citoplasma. La contrazione inizia
quando la concentrazione di calcio aumenta. Gli ioni calcio si legano alla calmodulina e attiva
l’enzima chinasi della catena leggera della miosina.
Alla base della testa della miosina si trova una piccola proteina di regolazione chiamata
catena leggera della miosina, ruolo importante nella regolazione della contrazione e del
rilasciamento. La fosforilazione e la defosforilazione della catena leggera della miosina
controllano la contrazione e il rilasciamento del muscolo liscio.
La fosforilazione della miosina aumenta l’attività ATPasica; quando essa è elevata, i legami
actina-miosina e il ciclo dei punti trasversali aumentano la tensione generata dal muscolo.
La defosforilazione della catena leggera della miosina da parte dell’enzima fosfatasi riduce
l’attività ATPasica della miosina. La defosforilazione della miosina non comporta
necessariamente il rilasciamento del muscolo. Infatti, può restare legata all’actina per un
certo tempo, si parla di stato di blocco della miosina. Questa condizione mantiene la tensione
della cellula muscolare con uno scarso consumo di ATP.
27. FUSI NEUROMUSCOLARI ED ORGANI DEL GOLGI: RUOLO NEI RIFLESSI MIOTATICO E DA
RILASCIAMENTO DEL MUSCOLO.
I riflessi sono innescati da uno stimolo che attiva un recettore sensoriale e possono essere
classificato secondo diversi criteri in base:
Alla sezione efferente del snc che controlla la risposta:
o Riflessi somatici, sono riflessi che coinvolgono i motoneuroni somatici e i muscoli
scheletrici
o Riflessi autonomici o viscerali, sono riflessi che presentano una risposta controllata
dai neuroni del sistema nervoso autonomi
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Alla sede del SNC in cui viene integrata la risposta:
o Riflessi spinali, integrati a livello del midollo spinale
o Riflessi corticali, integrati a livello encefalico
Al fatto che il riflesso sia innato o acquisito:
o Riflessi innati, riflessi geneticamente determinati
o Riflessi acquisiti
Al numero di numero di neuroni presenti nella via riflessa:
o Riflesso monosinaptico
o Riflesso polisinaptico
I riflessi autonomici sono definiti anche viscerali perché spesso coinvolgono gli organi interni.
Alcuni di questi riflessi sono spinali e sono modulati da informazioni inibitorie o eccitatorie di
origine encefalica, altri sono integrati a livello encefalico. Questi riflessi sono tutti
polisinaptici e molti hanno attività tonica.
I riflessi motori somatici sono coinvolti in quasi tutte le nostre azioni e sono costituite da:
▪ Recettori sensoriali, propriocettori che sono localizzati all’interno dei muscoli scheletrici,
nelle capsule articolari e nei legamenti. Rilevano la posizione delle diversi parti del corpo
nello spazio.
▪ Interneuroni eccitatori e inibitori, costituiscono vie attraverso le quali il SNC integra il
segnale
▪ Motoneuroni somatici, trasportano il segnale in uscita. Quelli che innervano le fibre
contrattili vengono definiti motoneuroni alpha.
▪ Fibre extrafusali, sono gli effettori cioè le fibre contrattili del muscolo scheletrico.
Nel muscolo ci sono 3 tipi di propriocettori:
• Recettori articolari: sono localizzati nelle capsule e nei legamenti di ogni articolazione e
portano l’informazione sulla posizione dei segmenti corporei.
• Fusi neuromuscolari: sono recettori sensibili allo stiramento del muscolo, cioè inviano
informazioni sulla lunghezza del muscolo e sulle sue variazioni. Presente in tutti i muscoli.
• Organi tendinei del Golgi: sono recettori che si trovano a livello della giunzione tra tendine
e fibre muscolari, disposte in serie rispetto ad esse. I riflessi tendinei del Golgi causano il
rilasciamento, cioè sono responsabili di un riflesso inverso a quello di stiramento. Le fibre
afferenti dall’organo del Golgi eccitano interneuroni inibitori nel midollo spinale. Questi
interneuroni inibiscono i motoneuroni alpha che innervano il muscolo e così la contrazione
diminuisce o termina. L’organo del Golgi previene contrazioni eccessive che potrebbero
danneggiare il muscolo.
28. CO – ATTIVAZIONE ALPHA – GAMMA NEL MUSCOLO PERCHÉ?
Quando i motoneuroni alfa scaricano potenziali d’azione sul muscolo questo si contrae e
diminuisce la tensione sul fuso. Contemporaneamente i motoneuroni gamma che innervano
le parti polari delle fibre intrafusali si attivano provocando la contrazione e l’accorciamento.
La contrazione delle parti polari stira quella centrale mantenendo attive le terminazioni
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
nervose sensoriali. Il fuso così resta attivo anche quando il muscolo si contrae. L’eccitazione
contemporanea dei motoneuroni alfa e gamma è definita co-attivazione alfa-gamma.
29. CUORE: STRUTTURA ANATOMICA (CAMERE E VALVOLE) E FUNZIONE NEL SISTEMA
CICOLATORIO.
Il cuore è diviso in 4 camere costituite da 2 atri e 2 ventricoli e ogni metà funziona come una
pompa indipendente. Il lato destro del cuore riceve sangue dai tessuti e lo invia ai polmoni
per l’ossigenazione. Il cuore sinistro riceve il sangue nuovamente ossigenato dai polmoni e lo
pompa ai tessuti in tutto l’organismo.
Il cuore si trova all’interno della gabbia toracica, sul mediastino medio, sul lato ventrale.
L’apice appuntito del cuore è in basso e poggia sul diaframma, la base è posta in alto dietro lo
sterno, tra i 2 polmoni. Il cuore è avvolto da una sacca membranosa resistente detta
pericardio, all’interno del quale vi è un liquido pericardico che lubrifica la superficie esterna
del cuore. La parete del cuore è composta da 3 strati:
1. L’endocardio è il sottile strato epiteliale che riveste le cavità interne e forma le
valvole;
2. Il miocardio è lo stato muscolare, forma la struttura vera e propria della parete ed è
rinforzato internamente da uno strato di connettivo fibroso denso;
3. L’epicardio è una sottile membrana sierosa che riveste esternamente il cuore.
Sulla superficie dei ventricoli si possono osservare due solchi che contengono le arterie e le
vene coronarie che forniscono sangue al miocardio. Il sangue fluisce dalle vene negli atri e da
lì attraverso le valvole unidirezionali nei ventricoli che costituiscono le camere di pompaggio.
Esso lascia il cuore attraverso il tronco polmonare dal ventricolo destro e tramite l’aorta dal
ventricolo sinistro. Una seconda serie di valvole è situata alle uscite dei ventricoli in modo che
il sangue non possa refluire nel cuore una volta che è stato eiettato. La prima serie di valvole
è detta atrioventricolare (tra atrio e ventricolo) la seconda semilunare (sono poste tra i
ventricoli e le arterie). La valvola AV è formata da dei lembi sottili di tessuto unito alla base
da un anello di tessuto connettivo. I lembi sono connessi al lato del ventricolare attraverso
tendini ricchi di collagene. Le estremità opposte delle corde sono inserite sui muscoli
papillari. Le valvole AV non sono uguali tra di loro: quella di destra è dotata di tre lembi ed è
chiamata tricuspide, quella di sinistra invece possiede solo due lembi ed è detta bicuspide o
mitrale. Le valvole semilunari separano i ventricoli dai vasi principali. In particolare, la valvola
polmonare si trova tra il ventricolo destro ed il tronco polmonare, mentre quella aortica si
trova tra il ventricolo sinistro e l’aorta.
30. STRUTTURA DELLE CELLULE CONTRATTILI CARDIACHE E GRAFICO DEL LORO POTENZIALE
D’AZIONE.
La maggior parte del muscolo cardiaco è contrattile ma solo l’1% delle cellule miocardiche è
specializzato nel generare spontaneamente potenziale d’azione. Il cuore può contrarsi senza
una connessione con altre parti del corpo perché il segnale per la contrazione è miogeno (si
origina all’interno del muscolo stesso). Lo stimolo viene da cellule miocardiche specializzate
chiamate autoritmiche, dette anche pacemaker perché regolano la frequenza del battito
cardiaco. Queste cellule non presentano sarcomeri organizzati, sono più piccole ed hanno
poche fibre contrattili. Queste cellule si connettono tra loro tramite i dischi intercalari
costituiti da 2 tipi di giunzioni: i desmosomi che legano insieme cellule adiacenti e
trasferiscono la forza da una cellula all’altra e quelle comunicanti che connettono
elettricamente le cellule adiacenti. I tubuli T sono più grandi di quelle muscolari e il reticolo
sarcoplasmatico è più piccolo rispetto a quello scheletrico e infine i mitocondri occupano
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
circa un terzo del volume cellulare di una fibra contrattile cardiaca, infatti il muscolo cardiaco
consuma il 70-80% dell’ossigeno portato.
Il Ca2+ svolge un ruolo importante nella
generazione di potenziale elettrico. Nello specifico,
in queste cellule, la fase di depolarizzione rapida è
il risultato dell’ingresso di Na, e la successiva
ripolarizzazione è dovuta al k+ che esce dalla
cellula.
FASE 4: potenziale di membrana a riposo. Le
cellule contrattili miocardiche hanno un potenziale
di riposo stabile di circa -90 mV.
FASE 0: depolarizzazione. Quando un’onda di
depolarizzazione diffonde in queste cellule
attraverso i canali Na voltaggio-dipendenti si
aprono permettendo al sodio di entrare nella
cellula e depolarizzare rapidamente. In questa il
potenziale raggiunge circa i +20 mV prima che i
canali si chiudano.
FASE 1: ripolarizzazione iniziale. Quando i canali
del sodio si chiudono la cellula comincia a
ripolarizzarsi mentre il K esce attraverso i suoi
canali aperti.
FASE 2: il plateau. Il potenziale d’azione
raggiunge poi un plateau come risultato di due
eventi: la diminuzione della permeabilità del K e
l’aumento della permeabilità al Ca2+. I canali del
calcio si sono attivati alla depolarizzazione e si
sono aperti durante le fasi 0 e 1, quindi il calcio
entra nella cellula. Allo stesso tempo i canali del K
si chiudono. Questa combinazione fa sì che il
potenziale d’azione raggiunga un plateau.
FASE 3: ripolarizzazione rapida. Il plateau termina
quando i canali calcio si chiudono e la
permeabilità al potassio aumenta. I canali "lenti"
del K+ responsabili di questa fase sono simili a
quelli nei neuroni: essi sono attivati da una
depolarizzazione, ma hanno una cinetica di apertura lenta. Quando questi canali lenti si
aprono, il K+ esce velocemente, riportando la cellula al suo potenziale di riposo.
31. SISTEMA DI CONDUZIONE DEL CUORE E POTENZIALE DELLE CELLULE PACEMAKER.
Le fasi del sistema di conduzione:
le fibre dell’atrio hanno un ritmo di 60 contrazioni al minuto, quelle del ventricolo 20. È
quindi necessaria una regolazione di questa attività per garantire un ritmo efficace.
Il sistema di conduzione è costituito da:
- Nodo seno – atriale
- Nodo atrio – ventricolare
- Fascio atrio – ventricolare (fascio di His)
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Fase 0
Il nodo SA invia il primo impulso (per quest’azione di innesco il nodo SA è considerato il
“pacemaker” naturale del cuore). L’impulso si propaga alle fibre cardiache degli atri; le cellule
del miocardio contrattile sono costituite da una struttura detta SINCIZIO FUNZIONALE; le
cellule sono collegate tra loro dai desmosomi e dai gap junction (quindi collegate sia
meccanicamente che elettricamente) e questo fa in modo che quando una cellula si contrae
trasmette la contrazione a tutte le cellule adiacenti.
Inoltre, vi è lo SCHELETRO CARDIACO: area che accoglie le valvole fatto di fibrocartilagine, la
quale isola elettricamente gli atri dai ventricoli e per tale motivo non si contraggono insieme
(tranne in un piccolo punto per permette il passaggio rallentato dell’impulso, in modo tale
che ci sia tempo per far completare le singole azioni degli atri e dei ventricoli).
Dal nodo SA l’impulso si propaga lungo le VIE INTERNODALI e giunge con alcuni centesimi di
secondo in ritardo al nodo AV (ritardo fondamentale per consentire agli atri di spingere il
sangue nei ventricoli prima che questi si contraggono) situato vicino alla giunzione tra atrio e
ventricolo destro.
Dal nodo AV parte il FASCIO di HIS che attraversa lo scheletro cardiaco e si porta all’interno
del setto interventricolare fino all’apice. Il FASCIO di HIS si divide in 2 rami: uno di destra e
uno di sinistra. Il ramo fa contrarre direttamente i 3 muscoli papillari di destra che tengono le
corde tendinee e mantengono chiuse le valvole.
Il ramo di sinistra fa contrarre i 2 muscoli papillari del rispettivo lato e un gruppo di cellule
chiamate del PURKINJE all’apice. Queste cellule sono in tutto il ventricolo di sinistra e in
minima parte in quello di destra. Queste cellule sono altamente eccitabili e quindi fanno in
modo che il ventricolo si contragga con più forza.
Le cellule pacemaker hanno la capacità unica di generare spontaneamente potenziale d’azione
senza ricevere stimoli dal sistema nervoso, questa capacità deriva appunto dal loro potenziale di
membrana instabile. Infatti, si pensa che le cellule autoritmiche contengano canali diversi da quelli
degli altri tessuti eccitabili. Quando il potenziale della membrana cellulare è -60 mV, si aprono i
canali If che sono permeabili sia al K+ sia al Na+ Questi canali sono chiamati canali If, perché
permettono alla corrente (l) di fluire con una cinetica particolare. Questo potenziale di membrana
instabile è detto potenziale pacemaker piuttosto che potenziale di membrana a riposo, perché
esso non si trova mai a un valore costante. Quando i canali If si aprono per potenziali di membrana
negativi, il Na che entra supera il K che esce. L’ingresso netto di cariche positive depolarizza
lentamente la cellula auto-ritmica. Nel momento in cui il potenziale di membrana diventa più
positivo, i canali If, si chiudono gradualmente e alcuni canali del Ca2+ si aprono. Il conseguente
ingresso di Ca2+ continua la depolarizzazione, e il potenziale di membrana si sposta
progressivamente verso il valore soglia. Quando il potenziale di membrana raggiunge la soglia, si
aprono ulteriori canali del Ca2+. Il Ca2+ entra velocemente nella cellula, generando la fase di
depolarizzazione rapida del potenziale di azione. Quando i canali del Ca 2+ si chiudono al picco del
potenziale d'azione, i canali lenti del K+ si aprono ne consegue l'efflusso di K+ che risulta quindi
responsabile della fase di ripolarizzazione del potenziale d'azione auto-ritmico. La velocità alla
quale le cellule pacemaker si depolarizzano determina la frequenza alla quale il cuore si contrae
(frequenza cardiaca).
(grafico pagina successiva)
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
32. MODULAZIONE DELLA FREQUENZA CARDIACA DA PARTE DEL SISTEMA NERVOSO
AUTONOMO.
La frequenza cardiaca media del cuore in un adulto a riposo è di circa 70 bpm (battiti per
minuto). Atleti allenati possono avere una frequenza di 50 bpm o meno, soggetti ansiosi
anche di circa 125 bpm e infine i bambini hanno mediamente frequenze cardiache più
elevate degli adulti. La frequenza cardiaca è influenzata dal SNA attraverso il controllo
antagonista, in particolare rallentandola tramite un’attività parasimpatica o aumentandola
attraverso un’attività simpatica.
Controllo Parasimpatico: il neurotrasmettitore parasimpatico acetilcolina rallenta la
frequenza cardiaca poiché attiva dei recettori colinergici muscarinici che influenzano i canali
di potassio e calcio della cellula pacemaker. La permeabilità al potassio aumenta quindi la
cellula si iperpolarizza in modo che il potenziale pacemaker inizio ad un valore soglia più
negativo, contemporaneamente la permeabilità al calcio dimunuisce. Questa riduzione
influisce sulla velocità di depolarizzazione e questi 2 effetti fanno sì che la cellula impieghi più
tempo a raggiungere la soglia ritardando così la frequenza cardiaca.
Controllo Simpatico: la noradrenalina e adrenalina aumentano il flusso ionico sia attraverso i
canali If sia attraverso i canali di calcio. L’ingresso più rapido di cationi accelera la frequenza di
depolarizzazione delle cellule pacemaker così che la cellula raggiunga il livello soglia più
velocemente. Quando il messaggero AMPc si lega per aprire i canali If essi rimangono aperti
più a lungo. L’aumento di permeabilità agli ioni sodio e calcio durante la fase pacemaker
accelera la depolarizzazione e la frequenza cardiaca.
33. LE 5 FASI DEL CICLO CARDIACO.
Un ciclo cardiaco comprende tutti gli eventi associati al flusso del sangue attraverso il cuore
durante un battito cardiaco.
• Il cuore a riposo: diastole atriale e ventricolare. Fase in cui sia gli atri che i
ventricoli sono rilasciati.
Gli atri si stanno riempiendo di sangue proveniente dalle vene, i ventricoli hanno
appena completato la contrazione. Quando i ventricoli si rilasciano le AV si aprono
e il sangue fluisce per gradiente di pressione dagli atri ai ventricoli. L’80% del
sangue che entrerà nei ventricoli arriva in questa fase.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
• Completamento del riempimento ventricolare: sistole atriale. In questa fase si
completa il restante 20% di riempimento del sangue nei ventricoli.
La sistole atriale inizia in seguito all’onda di depolarizzazione che invade gli atri.
L’aumento di pressione che accompagna la contrazione spinge il sangue nei
ventricoli. Una piccola quantità di sangue viene spinta indietro nelle vene perché
non esistono valvole per bloccare il reflusso.
• Fase iniziale della contrazione ventricolare e primo tono cardiaco.
Mentre gli atri si contraggono l’onda di depolarizzazione si muove lentamente
attraverso le cellule di conduzione nel nodo AV, poi rapidamente lungo le fibre del
Purkinje fino all’apice del cuore. La sistole (contrazione) ventricolare inizia a
questo punto, il sangue forza la chiusura delle valvole AV e le vibrazioni derivanti
dalla chiusura delle valvole genera il primo tono cardiaco S1. Con entrambe le serie
di valvole AV chiuse il sangue resta bloccato nei ventricoli che però continuano a
contrarsi.
Questa fase è detta contrazione ventricolare isovolumetrica per sottolineare il
fatto che il volume di sangue non cambia.
• Il cuore come pompa: eiezione ventricolare.
Quando i ventricoli si contraggono generano una pressione sufficiente ad aprire le
valvole semilunari e spingere il sangue nelle arterie. La pressione generata dalla
contrazione ventricolare diventa la forza motrice del flusso ematico. Il ventricolo
manda fuori il 50% di sangue.
Durante questa fase le valvole AV restano chiuse e gli atri continuano a riempirsi.
• Rilasciamento ventricolare e secondo tono cardiaco.
Alla fine dell’eiezione ventricolare, i ventricoli iniziano a rilasciarsi e di
conseguenza la pressione ventricolare diminuisce determinando il reflusso del
sangue verso il cuore con la conseguente chiusura delle valvole semilunari. Le
vibrazioni generate dalle valvole semilunari che si chiudono costituisce il secondo
tono cardiaco S2.
Quando si chiudono le valvole semilunari, anche le valvole AV restano chiuse
perché la pressione ventricolare, anche se diminuita, resta maggiore di quella
atriale. Questo periodo è detto rilasciamento ventricolare isovolumetrico.
Quando la pressione ventricolare scende al di sotto di quella atriale le valvole AV
si aprono, il sangue comincia a fluire nei ventricoli e ricomincia un altro ciclo
cardiaco.
34. IL CICLO CARDIACO SPIEGATO ATTRAVERSO LA CURVA PRESSIONE-VOLUME DEL VENTRICOLO
SINISTRO.
Il flusso di sangue nel cuore si sposta da zone a pressione più elevata a zone a pressione più
bassa. Quando il cuore si contrae, la pressione aumenta e il sangue esce da esso verso zone a
pressione minore.
PUNTO A: qui inizia il ciclo. Il ventricolo ha completato la contrazione e contiene unna
quantità minima di sangue che manterrà durante l’intero ciclo. Il sangue sta fluendo nell’atrio
dalle vene polmonari. Quando la pressione nell’atrio supera quella esistente nel ventricolo la
valvola mitrale si apre; di conseguenza, il sangue entrerà nel ventricolo aumentandone il
volume (A → B).
PUNTO B: il ventricolo contiene il massimo volume sanguigno, chiamato volume
telediastolico. Individuo di circa 70 kg a riposo il volume è di circa 135 ml. Durante il
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
passaggio dal punto B al punto C (A → B), il ventricolo continua a contrarsi causando un
rapido aumento della pressione in questa camera.
PUNTO C: appena la pressione ventricolare supera quella esistente nell’aorta, la valvola
aortica si apre. La pressione continua ad aumentare man mano che il ventricolo si contrae
ulteriormente, mentre il volume ventricolare diminuisce poiché il sangue è spinto fuori dal
cuore (C → D).
PUNTO D: il cuore non si svuota completamente ogni volta che il ventricolo si contrae. La
quantità di sangue residua nel ventricolo alla fine della contrazione è nota come volume
telesistolico (valore medio di 65 ml). Esso rappresenta la quantità minima di sangue che il
ventricolo contiene durante un ciclo cardiaco.
Riassumendo:
A → il ciclo inizia, si apre la valvola
mitrale.
AB → riempimento ventricolare
(aumento di volume) +35 ml
(volume telediastolico)
B → chiusura valvola mitrale
BC → fase di contrazione atriale
C → si apre la valvola semilunare
aortica
CD → il sangue viene mandato in
circolo, la massima forza espulsiva il
cuore la raggiunge un po’ prima di
raggiungere il punto D
D → chiusura valvola semilunare
aortica (volume telesistolico)
DA → rilasciamento ventricolare
isovolumetrico.
35. LA GITTATA CARDIACA: COME SI CALCOLA E COME AUMENTA DURANTE L’ESERCIZIO.
La gittata cardiaca (GC) è il volume di sangue pompato da un ventricolo in un minuto. La
gittata cardiaca è un indicatore del flusso ematico totale di tutto l’organismo.
GC = Frequenza cardiaca X Gittata sistolica
GC = 72 battiti/min X 70 ml/min = 5040 ml/min (circa 5 l/min)
Gittata sistolica è la quantità di sangue pompata da un ventricolo durante una contrazione.
Gittata sistolica = Vol. di sangue prima della contrazione – Vol. di sangue dopo la
contrazione
Gittata sistolica = Volume telediastolico – Volume telesistolico
Gittata sistolica = 135 ml – 65 ml = 70 ml
A riposo la gittata cardiaca è di circa 5 l/min; sotto sforzo può aumentare fino a 15-20 l/min.
in un soggetto allenato, durante l’esercizio fisico, può aumentare fino a 30-35 l/min.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
36. LEGGE DI FRANK – STARLING E L’IMPORTANZA DEL PRECARICO NEL DETERMINARE LA
GITTATA SISTOLICA.
Quando una quantità ulteriore di sangue entra nel cuore, il cuore si contrae con maggiore
forza ed espelle più sangue. Questa relazione è nota come legge di Frank – Sterling.
Secondo questa legge, la gittata sistolica aumenta quando il volume telediastolico aumenta. Il
volume telediastolico normalmente è determinato dal ritorno venoso, che a sua volta è
influenzato da 3 fattori:
• Contrazione o compressione delle vene che riportano il sangue al cuore. Questa
contrazione è dovuta all’azione della pompa muscolare scheletrica. Durante
l’esercizio fisico, che coinvolge gli arti inferiori, la pompa muscolare scheletrica
facilita il ritorno venoso al cuore.
• Variazione di pressione nell’addome e nel torace durante la respirazione. Azione
della pompa respiratoria. Quando la gabbia toracica si espande, il diaframma si
abbassa verso l’addome allargando la cavità toracica provocando una riduzione di
pressione. Questa bassa pressione diminuisce la pressione nella vena cava
inferiore nel suo passaggio attraverso il torace facendo sì che una maggiore
quantità di sangue sia richiamata dalle vene addominali. Questa combinazione di
aumento della pressione nelle vene addominali e diminuzione in quelle toraciche
aumenta il ritorno venoso durante l’inspirazione.
• Innervazione simpatica delle vene. Permette all’organismo di ridistribuire il
sangue venoso nel distretto arterioso della circolazione.
37. L’ELETTROCARDIOGRAMMA: GRAFICO ED INTERPRETAZIONE.
È possibile utilizzare elettrodi di superficie per registrare l’attività elettrica interna, perché le
soluzioni saline, come il nostro liquido extracellulare a base di NaCl, sono dei buoni
conduttori di elettricità.
L’ECG è una registrazione extracellulare che rappresenta
la somma dei potenziali d’azione che hanno luogo nel miocardio.
In un normale ECG si osservano tre onde principali:
• Onda P. la prima onda e corrisponde alla
depolarizzazione degli atri.
• Complesso QRS. Trio di onde che rappresenta
l’onda progressiva della depolarizzazione
ventricolare. Incorpora la ripolarizzazione
atriale.
• Onda T. Onda finale e rappresenta la
ripolarizzazione dei ventricoli.
Un ECG fornisce informazioni sulla frequenza cardiaca e sul ritmo cardiaco, sulla velocità di
conduzione e, persino, sulle condizioni dei tessuti del cuore.
38. RUOLO DI VASI E POMPA MUSCOLATURA SCHELETRICA NELL’AIUTARE IL CUORE A POMPARE
IL SANGUE.
La pressione prodotta dalla contrazione del ventricolo sinistro è immagazzinata nelle pareti
elastiche delle arterie e rilasciata lentamente attraverso il loro ritorno elastico. Le arterie
sono note come riserva di pressione del sistema circolatorio. Le vene invece agiscono come
riserva di volume dalla quale, se la pressione sanguigna scende troppo, il sangue può essere
inviato alla parte arteriosa della circolazione. Le pareti dei vasi sanguigni sono composte da
strati di muscolatura liscia, tessuto connettivo elastico e tessuto connettivo fibroso. Il
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
rivestimento interno di tutti i vasi sanguigni è costituito da un sottile strato di endotelio
organizzato in strati circolari o a spirale. La vasocostrizione determina una riduzione del
diametro del lume dei vasi, mentre la vasodilatazione un aumento. La contrazione di questa
muscolatura dipende anch’essa dall’ingresso del calcio attraverso i propri canali ma anhe
dall’ingresso di ormoni e sostanze paracrine. La muscolatura liscia comincia a comparire nelle
pareti delle venule grandi. Il sangue delle venule di solito fluisce nelle vene che aumentando
di diametro andranno a formare vene più grandi chiamate vene cave che si svuoteranno
nell’atrio destro. Le vene sono più numerose delle arterie e hanno un diametro maggiore
poiché contengono più della metà di tutto il sangue del sistema circolatorio. Le vene sono
localizzate in una zona più in superficie del corpo rispetto alle arterie, inoltre hanno pareti più
sottili con meno tessuto elastico e di conseguenza si dilatano più facilmente quando si
riempiono di sangue.
L’aorta e le arterie principali sono caratterizzate da pareti sia rigide che elastiche; per questo
è necessaria una grande quantità di energia per distendere le loro pareti. Le arterie e le
arteriole sono caratterizzate da percorsi divergenti, dal momento che la maggior parte delle
arterie si divide in arterie sempre più piccole, le caratteristiche delle pareti cambiano
diventando più elastiche e più ricche di muscolatura.
39. PRESSIONE ARTERIOSA E SANGUIGNA: SUA MISURAZIONE E COME CAMBIA LUNGO LA
CIRCOLAZIONE.
La pressione arteriosa riflette l’andamento della pressione generata dalla pompa cardiaca e
dal momento che questa è pulsatile. La indichiamo come Pressione Arteriosa Media (PAM).
𝟏
PAM = Pdiastolica + (Psistolica – Pdiastolica)
𝟑
1
PAM = 80 mmHg + 3 (120 – 80 mmHg) = 93 mmHg
La pressione arteriosa media è la forza propulsiva del flusso sanguigno e anche un equilibrio
tra il flusso sanguigno che entra nelle arterie e quello che esce.
Dipende da tre fattori:
• La frequenza cardiaca. La PAM dovrebbe aumentare in seguito ad un
innalzamento della frequenza cardiaca e del volume di eiezione.
• Il volume di eiezione ventricolare. Quando la PAM è stazionaria il sangue entra
nell’arteria alla stessa velocità con la quale esce.
• La resistenza periferica totale. Quando la PAM aumenta in seguito ad un
improvviso aumento della gittata, il sangue entrerà più velocemente di quanto ne
esce, il volume di sangue nell’arteria aumenta e il vaso si espande.
La pressione arteriosa si misura tramite uno sfigmomanometro. Il manicotto circonda la parte
superiore del braccio e viene gonfiato fino a quando non esercita una pressione più alta di
quella sistolica. A questo punto la pressione del manicotto viene gradualmente ridotta e
quando questa scende al di sotto della pressione arteriosa sistolica il sangue ricomincia a
fluire; il sangue cerca di fluire nell’arteria ancora compressa creando un flusso turbolento e
grazie allo stetoscopio si riesce a percepire, sull’arteria brachiale, un rumore chiamato tono di
Korotkoff. Quando il manicotto non comprime più l’arteria ritorna il flusso laminare e il
rumore scompare.
La pressione alla quale il tono Korotkoff è udibile per la prima volta rappresenta la pressione
più alta e corrisponde alla pressione sistolica. Il punto in cui il tono di Korotkoff scompare è la
pressione più bassa dell’arteria e corrisponde alla pressione diastolica.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
40. REGOLAZIONE DELLA PRESSIONE ARTERIOSA E RIFLESSO BAROCETTIVO.
Il SNC coordina il controllo riflesso della pressione arteriosa e la distribuzione del sangue ai
tessuti. Il principale centro di integrazione si trova nel bulbo, centro di controllo
cardiovascolare (CCCV). La sua funzione principale è di garantire un adeguato flusso
sanguigno a cervello e cuore attraverso il mantenimento di una PAM sufficiente.
Il riflesso più importante del controllo omeostatico della pressione arteriosa è il riflesso
barocettivo. All’interno delle arterie carotidi e dell’aorta sono localizzati dei meccanocettori
sensibili allo stiramento, i barocettori, questi monitorano la pressione del sangue che fluisce
al cervello (barocettori carotidei) e al corpo (barocettori aortici). I barocettori sono recettori
di stiramento tonicamente attivi che producono continuamente potenziali d’azione. Quando
l’aumento della pressione arteriosa provoca lo stiramento della membrana delle terminazioni
nervose dei barocettori, la frequenza di scarica aumenta; se la pressione scende la frequenza
di scarica diminuisce. Quindi, il CCCV integra l’informazione sensoriale e innesca una risposta
appropriata. La risposta barocettiva è abbastanza rapida è modifica (1) gittata cardiaca e (2)
resistenza periferica fino a portarla al valore normale entro due battiti cardiaci dallo stimolo.
I segnali d’uscita (efferenti) del CCCV sono condotti sia da neuroni simpatici sia parasimpatici
del SNA.
La resistenza periferica è sotto il controllo tonico del simpatico e l’aumento di scarica
determina vasocostrizione. L’aumento dell’attività simpatica aumenta la frequenza cardiaca
aumentando la scarica del nodo SA, diminuisce il tempo di conduzione atrio – ventricolare e
aumenta la forza di contrazione del miocardio. L’aumento dell’attività parasimpatica riduce la
frequenza cardiaca.
Quando la pressione arteriosa aumenta, aumenta la frequenza di scarica dei barocettori
diretta al CCCV bulbare che innesca una risposta, che consiste nell’aumento dell’attività
parasimpatica e nella diminuzione dell’attività simpatica per ridurre la frequenza cardiaca e
dilatare le arteriole.
Quando la frequenza cardiaca diminuisce, la diminuzione dell’attività simpatica causa la
dilatazione delle arteriole abbassando la loro resistenza e permettendo ad una quantità
maggiore di sangue di lasciare le arterie.
Il riflesso barocettivo è sempre in funzione. Una variazione della pressione può tradursi in un
cambiamento sia della gittata cardiaca sia della resistenza periferica o nella variazione di una
sola delle due variabili.
41. IL SANGUE: COMPOSIZIONE E FUNZIONE DELLE SUE COMPONENTI CORPUSCOLATE.
Il sangue è responsabile del trasporto delle sostanze tra i vari distretti del cuore. Il volume
totale di sangue in un soggetto corrisponde a circa il 7% del suo peso. Nel sangue ci sono
anche elementi corpuscolari e plasma.
Il plasma è la porzione liquida del sangue in cui sono sospesi gli elementi corpuscolati. La
componente principale è l’acqua e corrisponde a circa il 92%, le proteine ne rappresentano
un altro 7% e il restante 1% è costituito da altre molecole organiche disciolte. Le proteine
albumine, le globuline e le proteine della coagulazione rappresentano circa il 90% delle
proteine plasmatiche. La presenza di proteine nel plasma rende la pressione osmotica del
sangue maggiore di quella del liquido interstiziale e partecipano a molte funzioni, tra la
coagulazione e la difesa contro agenti patogeni esterni e fungono da trasportatori.
I tre principali elementi corpuscolati presenti nel sangue sono:
• Globuli rossi (o eritrociti)
• Globuli bianchi (o leucociti)
• Le piastrine
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Solo i globuli bianchi sono cellule complete, perché i globuli rossi perdono il nucleo e le
piastrine sono frammenti di una cellula parentale gigante – “il megacariocita”.
I globuli rossi sono gli elementi corpuscolati del sangue più abbondanti. La loro funzione
principale è trasportare ossigeno dai polmoni alle cellule e anidride carbonica dalle cellule ai
polmoni. Non presentano nucleo, reticolo endoplasmatico e mitocondri. Essi sono molto
flessibili utile a modificare la propria forma per passare attraverso i capillari dell’apparato
circolatorio.
Le piastrine sono essenziali per la coagulazione, il processo grazie al quale il sangue si
solidifica bloccando l’emorragia dai vasi danneggiati. Il processo che permette di mantenere il
sangue all’interno dei vasi è detto emostasi e si articola in tre stadi: (1) vasocostrizione –
riduce temporaneamente il flusso e la pressione del sangue; (2) formazione del tappo
piastrinico che va a bloccare la lesione. Inizia con l’adesione piastrinica quando esse
aderiscono al collagene esposto nella parete danneggiata. Il collagene esposto e un fattore
tissutale innescano il terzo passaggio; (3) coagulazione del sangue.
I globuli bianchi hanno un ruolo fondamentale nella risposta immunitaria dell’organismo.
I diversi elementi corpuscolati discendono da un’unica cellula staminale emopoietica
pluripotente presente principalmente nel midollo osseo. Il processo in vengono prodotti
questi elementi si chiama emopoiesi.
La durata della vita dei globuli bianchi è notevolmente più corta rispetto a quella dei globuli
rossi. Infatti, i globuli bianchi hanno un’emivita di sei ore circa, mentre i globuli rossi vivono
per circa quattro mesi.
42. APPARATO RESPIRATORIO: STRUTTURA E FUNZIONE DI POLMONI, ALVEOLI E GABBIA
TORACICA.
Le funzioni principali dell’apparato respiratorio sono:
• Scambio di gas tra atmosfera e sangue.
• Regolazione omeostatica del pH corporeo.
• Protezione da patogeni inalati o da sostanze irritanti.
• Vocalizzazione.
Il sistema respiratorio è costituito:
• Sistema di conduzione o vie aree. Va dall’ambiente esterno alla superficie di
scambio dei polmoni;
• Gli alveoli. Una serie di strutture semisferiche interconnesse associate ai capillari
alveolari e formano la superficie di scambio attraverso cui l’ossigeno diffonde
dall’aria inspirata al sangue e l’anidride carbonica diffonde dal sangue all’aria che
verrà espirata;
• Ossa e muscoli del torace e addome. Permettono la ventilazione.
Il sistema respiratorio è suddiviso in due parti: tratto respiratorio superiore che comprende
bocca, cavità nasali, faringe, laringe e trachea; tratto respiratorio inferiore che comprende
trachea, due 2 bronchi primari con le loro ramificazioni e dai polmoni.
(1) I polmoni sono costituiti da tessuto leggero e spugnoso. Questi organi a forma di cono
irregolare occupano la cavità toracica e hanno la base poggiata sul diaframma. I bronchi
collegano i polmoni alla trachea. La pleura è costituita da diversi strati di tessuto
connettivo elastico e da numerosi capillari.
(2) Gli alveoli raggruppati alle estremità dei bronchioli terminali costituiscono la maggior
parte del tessuto polmonare. La funzione primaria degli alveoli è lo scambio di gas tra
l’aria che contengono e il sangue. Ogni minuscolo alveolo è costituito da un singolo strato
sottile di scambio. Le cellule alveolari di II tipo, più piccola ma più spesse, sintetizzano e
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
secernano una sostanza chimica chiamata surfactante. Questa sostanza si mescola con il
sottile strato di liquido che riveste gli alveoli per facilitare l’espansione polmonare
durante la ventilazione. Le cellule di II tipo contribuiscono anche a minimizzare il volume
di liquido presente negli alveoli trasportando soluti, seguiti da acqua fuori dagli spazi
areai alveolari. Le cellule alveolari di tipo I sono più grandi, occupano il 95% della
superficie alveolare e sono così sottili che i gas possono diffondere rapidamente
attraverso di esse. Il tessuto connettivale presente ha molto fibre di elastina e collagene
che permettono un ritorno elastico del tessuto polmonare.
(3) Il torace è delimitato dalle ossa della colonna vertebrale, dalle coste e dai muscoli
associate ad esse. Le coste e la colona vertebrale formano le pareti laterali superiori della
cassa toracica. Due gruppi di muscoli intercostali (interni ed esterni) connettono le 12
paia di coste. I muscoli sternocleidomastoidei e i muscoli scaleni si portano dalla testa e
dal collo fino allo sterno e alle prime due coste. Dal punto di vista funzionale, la cassa
toracica è un contenitore sigillato che al suo interno ha tre sacchi membranosi.
43. LE 4 FASI DELLA RESPIRAZIONE ESTERNA: QUALI SONO E QUALI COMPITI SVOLGONO?
Le 4 fasi della respirazione esterna sono:
• La ventilazione. È il processo in cui si ha lo scambio di aria tra atmosfera e
polmoni, questo processo è costituito da una prima fase inspiratoria e da una
seconda fase espiratoria.
• Lo scambio gassoso polmonare. (polmone → sangue)
• Il trasporto sanguigno
• Lo scambio gassoso sanguigno. (sangue → tessuti)
44. LA VENTILAZIONE: MUSCOLI IMPLICATI E MECCANICA RESPIRATORIA.
La ventilazione è il flusso di massa che scambia l’aria fra l’atmosfera e gli alveoli. Un singolo
ciclo respiratorio è costituito da una fase di inspirazione che determina: (1) aumento del
volume toracico, (2) abbassamento della pressione intrapolmonare, (3) ingresso d’aria;
seguita da una fase di espirazione la quale determina: (1) diminuzione del volume toracico,
(2) innalzamento della pressione intrapolmonare, (3) fuoriuscita d’aria.
I muscoli coinvolti nella fase inspiratoria sono:
• Diaframma
• Intercostali esterni
• Scaleni
• Sternocleidomastoidei
I muscoli coinvolti nella fase espiratoria invece sono:
• Addominali
• Intercostali interni
Nella respirazione a riposo, nella fase inspiratoria il 70% del lavoro inspiratorio è compiuto
dal diaframma e il restante 30% dai muscoli intercostali esterni. L’espansione della gabbia
toracica è causata dal movimento del diaframma verso il basso, delle coste verso l’alto e dello
sterno in avanti.
Alla fine della fase inspiratoria i muscoli coinvolti si rilasciano, il ritorno del diaframma e delle
coste alla posizione di partenza è dovuto al ritorno elastico dei polmoni. Poiché l’espirazione
è dovuta ad un ritorno elastico passivo viene definita espirazione passiva.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
45. COME CAMBIANO PRESSIONE E VOLUME INTRAPOLMONARE, E PRESSIONE INTRA-PLEURICA
DURANTE LA VENTILAZIONE A RIPOSO.
La legge di Boyle afferma che se si riduce il volume di un gas la pressione aumenta. Se il
volume aumenta la pressione diminuisce.
Quando il diaframma si contrae (nella ventilazione tranquilla) aumenta il volume della cavità
toracica. La contrazione del diaframma determina tra il 60-70% della variazione del volume
polmonare. Il movimento della cassa toracica determina il restante 25-40%. La combinazione
di questi due movimenti allarga la cassa toracica in tutte le direzioni. Quando il volume della
cavità toracica aumenta, la pressione all’interno diminuisce e l’aria fluisce nei polmoni.
TEMPO 0: tra un atto respiratorio e l’altro,
la pressione alveolare è uguale a quella
atmosferica. In assenza di un gradiente di
pressione non si ha un flusso d’aria.
TEMPO 0-2s: INSPIRAZIONE. Quando
l’inspirazione inizia i muscoli per questa
fase si contraggono e il volume toracico
aumenta. Con l’aumento del volume si
riduce la pressione intrapolmonare e l’aria
inizia a muoversi dentro gli alveoli. Mentre
l’aria continua ad entrare negli alveoli la
pressione aumenta gradualmente, fino a
quado la pressione all’interno dei polmoni
è uguale a quella atmosferica (punto A3).
Al termine dell’inspirazione la pressione
alveolare è uguale a quella atmosferica.
TEMPO 2-4s: ESPIRAZIONE. Mentre il
volume dei polmoni e del torace
diminuisce durante l’espirazione, la
pressione dell’aria nei polmoni aumenta.
La pressione alveolare ora è superiore a
quella atmosferica quindi il flusso d’aria si
inverte e l’aria esce dai polmoni.
TEMPO 4s: al termine dell’espirazione, il
movimento dell’aria si blocca quando la
pressione alveolare è di nuovo uguale a
quella atmosferica. A questo punto
termina il ciclo respiratorio e ne inizia un
altro.
La ventilazione richiede che i polmoni si muovano in associazione con la distensione e il
rilasciamento del torace. La forza di coesione esercitata dal liquido tra le due membrane
pleuriche provoca l’adesione dei polmoni alla cassa toracica. Le due membrane pleuriche
rimangono adese tramite il liquido pleurico, di conseguenza i polmoni si mantengono distesi
occupando l’intero volume della cassa toracica. Allo stesso tempo il ritorno elastico dei
polmoni generare una forza diretta dall’interno che tende ad allontanare i polmoni dalla
parete toracica. All’inizio dell’inspirazione la pressione intra pleurica è circa -3 mmHg (punto
B1) e mentre l’inspirazione procede il tessuto polmonare si oppone all’azione di allungamento
e i polmoni tendono ad allontanarsi dalla parete toracica portando la pressione intra pleurica
a valori ancora più negativi (punto B2). Quindi al termine di un’inspirazione, quando i polmoni
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
sono completamente espansi, la pressione intra pleurica scende attorno a -6 mmHg (punto
B2). Durante l’espirazione, invece, la cassa toracica torna alla propria posizione di partenza e
la pressione intra pleurica torna al proprio valore normale di circa -3mmHg (punto B3).
La maggior parte del movimento ventilatorio è utilizzato per vincere le resistenze
dell’elasticità polmonare e della cassa toracica all’estensione. La capacità dei polmoni di
espandersi è definita complianza. La complianza è diversa dall’elastanza. Il fatto che un
polmone si espanda facilmente – elevata complianza – non significa necessariamente che
riassuma il proprio volume a riposo quando la forza che tende a dilatarlo venga meno
(elastanza).
46. PLEURE: FUNZIONE ED IMPORTANZA PER LA VENTILAZIONE.
Ciascun polmone si trova all’interno di un sacco pleurico formato da due foglietti pleurici, di
cui uno riveste l’interno della cavità toracica e l’altro la superficie esterna del polmone. Le
pleure sono membrane costituite da diversi strati di tessuto connettivo elastico. I due
foglietti sono tenuti insieme dal liquido pleurico. Questo liquido ha le funzioni di: (1) creare
superficie umida e scivolosa in modo che i due foglietti membranosi possano scorrere l’uno
sull’altro durante il movimento dei polmoni e (2) mantenere i polmoni a stretto contatto con
la parete toracica.
47. MI PARLI DELLA SPIROMETRIA: VOLUMI E CAPACITÀ POLMONARI CHE CON ESSA POSSONO
ESSERE MISURATI.
Lo spirometro è uno strumento che misura il volume di aria spostato ad ogni atto
ventilatorio.
Quando un soggetto è connesso ad uno spirometro, gli vengono chiuse le narici così da
costituire un sistema chiuso. Quando
inspira si crea un flusso d’aria dallo
spirometro ai polmoni e la penna di
registrazione si alza; quando espira
l’aria fluisce dai polmoni allo
spirometro e la penna si abbassa.
La quantità di aria che viene mossa
durante la ventilazione può essere
divisa in quattro volumi polmonari:
• Volume corrente: 500 ml
• Volume di riserva
respiratoria: 3000 ml
• Volume di riserva
espiratoria: 1100 ml
• Volume residuo: 1200 ml
Il volume corrente (VC) è il volume di aria che si muove durante una singola inspirazione o
espirazione e il suo valore medio durante la ventilazione tranquilla è circa 500 ml. Il volume
aggiuntivo inspirato oltre al volume corrente è il volume di riserva inspiratoria (VRI) che è di
circa 3000 ml. Il volume di aria esalata forzatamente dopo la fine di un’espirazione tranquilla
è detto volume di riserva espiratoria (VRE) ed è di circa 1100 ml. Il volume di aria presente
dopo un’espirazione massimale vie detto volume residuo (VR) e non può essere misurato
direttamente dallo spirometro e pertanto viene solo stimato ed è di circa 1200 ml.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
La somma di due o più volumi polmonari è detta capacità. La capacità vitale (CV) è la somma
dei tre volumi misurabili (VRI, VRE, VC). Essa rappresenta il massimo volume di aria che può
essere volontariamente spostata dentro e fuori l’apparato respiratorio durante un ciclo di
inspirazione – espirazione. La somma della capacità vitale e del volume residuo costituisce la
capacità polmonare totale (CPT), ovvero la somma dei quattro volumi.
48. VENTILAZIONE POLMONARE ED ALVEOLARE: COME SI CALCOLANO E DIFFERENZE TRA DI
ESSE.
L’efficacia della ventilazione può essere stimata calcolando la ventilazione polmonare totale,
cioè il volume di aria spostato dentro e fuori dai polmoni ad ogni minuto.
Ventilazione polmonare totale = Frequenza ventilatoria × Volume corrente
Ventilazione polmonare totale = 12 atti/min × 500 ml/atto = 6000 ml/min = 6 L/min
La ventilazione polmonare totale rappresenta lo scambio fisico dell’aria tra l’organismo e
l’ambiente, ma non è un buon indice della quantità di aria che raggiunge la superficie di
scambio alveolare, perché una parte resta nelle vie aree di conduzione, poiché queste vie non
riescono a scambiare gas con il sangue vengono dette spazio morto anatomico (SMA), il quale
𝟏
corrisponde a circa del volume corrente (circa 150 ml).
𝟑
Un indice più preciso dell’efficacia della ventilazione è la ventilazione alveolare, cioè la
quantità di aria che raggiunge gli alveoli ogni minuto.
Ventilazione alveolare = Frequenza ventilatoria × (Volume corrente – SMA)
Ventilazione alveolare = 12 atti/min × (500-150 ml/atto) = 4200 ml/min
Solo 4,2 L raggiungono gli alveoli
La ventilazione alveolare può essere influenzata da variazioni della frequenza o profondità
della ventilazione. La ventilazione volontaria massima, che comporta atti ventilatori molto
profondi e rapidi, può aumentare la ventilazione polmonare totale fino a 170 L/min.
49. COME CAMBIANO LE PRESSIONI ALVEOLARI DI O2 E CO2 CON IPO- ED IPER-VENTILAZIONE?
Quando la ventilazione alveolare supera i
valori normali si tratta di iperventilazione,
dove i valori di PO2 e PCO2 alveolare
rispettivamente salgono e scendono
rimanendo per lo più uguali e vanno a
formare una curva fisiologica.
Nell’ipoventilazione, quando diminuisce l’aria
fresca che entra negli alveoli, i valori variano
di molto, PO2 diminuisce e PCO2 aumenta
esponenzialmente; in questo caso si parla di
curva teorica o patologica dato che i valori
rappresentati non sono normali e portano alla
morte.
Nonostante un cambiamento marcato della
ventilazione possa influenzare la pressione
parziale dei gas negli alveoli, le due pressioni
parziali alveolari si modificano poco durante la
ventilazione normale a riposo. La PO2 alveolare rimane ad un valore di circa 100 mmHg e
PCO2 a circa 40 mmHg.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
50. SCAMBIO GASSOSO ALVEOLARE E TISSUTALE DI O2 E CO2.
Quando l’aria raggiunge gli alveoli, ossigeno e anidride carbonica diffondono dallo spazio
areo alveolare al sangue. La PO2 alveolare normale a livello del mare è di circa 100 mmHg. La
PO2 del sangue venoso “deossigenato” che arriva ai polmoni è di 40 mmHg. Quindi, l’ossigeno
si muove secondo il proprio gradiente di pressione parziale dagli alveoli ai capillari.
Quando il sangue arterioso raggiunge i capillari il gradiente si inverte.
In una cellula in condizioni a riposo la PO2 intracellulare è di 40 mmHg. Il sangue arterioso che
arriva alle cellule ha PO2 di 100 mmHg. Poiché la PO2 è minore nelle cellule, l’ossigeno
diffonde secondo il proprio gradiente di pressione parziale dal plasma alle cellule.
Al contrario la PCO2 è maggiore nei tessuti a causa della produzione metabolica di CO2. La
PCO2 cellulare in un soggetto a riposo è di circa 46 mmHg a fronte di una PCO2 nel plasma di
40 mmHg. Il gradiente determina il trasporto netto di anidride carbonica dalle cellule ai
capillari. La diffusione raggiunge l’equilibrio e il sangue venoso sistemico ha una pCO2 media
di 46 mmHg. A livello dei capillari polmonari il processo si inverte.
51. TRASPORTO GASSOSO SANGUIGNO DI O2 E CO2.
Gli eritrociti hanno un ruolo fondamentale nel garantire
che il trasporto del gas tra i polmoni e le cellule sia
adeguato a soddisfare le esigenze cellulari. Senza
l’emoglobina dei globuli rossi, il sangue non sarebbe in
grado di trasportare ossigeno sufficiente a sostenere la
vita. L’ossigeno è trasportato nel sangue in due modi:
disciolto nel sangue oppure legato all’emoglobina.
L’ossigeno è poco solubile nelle soluzioni acquose e
meno del 2% di tutto l’ossigeno è disciolto nel sangue,
quindi il restante 98% è trasportato dall’emoglobina.
Il trasporto di gas nel sangue include la rimozione
dell’anidride carbonica. Essa è un prodotto di scarto
della respirazione ed è molto più solubile dell’ossigeno
nei liquidi del corpo umano. Nonostante ciò,
l’emoglobina ha un ruolo fondamentale anche nel
trasporto di anidride carbonica. Solo circa il 7% della
CO2 trasportata dal sangue venoso è disciolta, il
restante 93% diffonde nei globuli rossi, dove il 70% è
convertito in ione bicarbonato e il 23% si lega all’emoglobina. È importante la rimozione della
CO2 dall’organismo poiché un’elevata PCO2 causa un disturbo del pH detto acidosi, deprime
l’attività del SNC causando coma o morte.
La CO2 dopo esser disciolta nel plasma passa nei globuli rossi, dove si trova l’enzima anidrasi
carbonica che catalizzata la reazione trasforma in acido carbonico (HCO3) che a sua volta si
andrà a scindere in idrogeno e ione bicarbonato che funge da tampone per l’organismo
poiché alza il pH essendo una sostanza basica e poi c’è l’idrogeno che abbassa il pH poiché è
acido. Quindi gli idrogeni sono legati all’emoglobina che funge da tampone evitando
l’abbassamento del pH; se però la CO2 è molto elevata, l’emoglobina non riesce più a
tamponare e allora gli idrogeni si accumulano andando a formare l’acidosi respiratoria.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
52. EMOGLOBINA: STRUTTURA, RUOLO E CURVA DI DISSOCIAZIONE.
L’emoglobina è una
proteina che lega
l’ossigeno tramite un
legame reversibile, questa
è un tetramero con
quattro catene proteiche
globulari, ognuna avvolta
attorno ad un gruppo eme
contenente ferro. In una
molecola di emoglobina
sono presenti quattro
atomi di ferro.
L’emoglobina legata
all’ossigeno si chiama
ossiemoglobina.
L’ossigeno disciolto
diffonde fuori dai capillari
sistemici nelle cellule, e la
riduzione della PO2
plasmatica che ne deriva
altera l’equilibrio della
reazione di legame
ossigeno – emoglobina. La
quantità dipende da due fattori: la PO2 del plasma che circonda i globuli rossi e il numero di
potenziali siti di legame dell’emoglobina disponibili all’interno dei globuli rossi. La PO2 del
plasma è il fattore principale che determina la percentuale dei siti di legame disponibili
dell’emoglobina occupati dall’ossigeno.
La forma delle curve di saturazione dell’ossi–emoglobina (o curva di dissociazione) riflette le
proprietà della molecola di emoglobina e la sua affinità per l’ossigeno. Si può notare che a
livello normale di PO2 alveolare e arterioso (100 mmHg), il 98% dell’emoglobina è legata
all’ossigeno. L’emoglobina si carica di quasi tutta la quantità di ossigeno che è in grado di
legare. La curva risulta essere quasi piatta a livelli di PO2 maggiori di 100 mmHg. Variazioni
anche ampie di PO2 determineranno solo piccole variazioni della percentuale di saturazione.
L’emoglobina non raggiungerà mai il 100% della saturazione fino a quando la PO2 non
raggiunge circa 650 mmHg. Finché la PO2 negli alveoli rimane superiore a 60 mmHg,
l’emoglobina è saturata per più del 90% e mantiene il trasporto O2 a livelli quasi normali.
Qualsiasi fattore in grado di cambiare la configurazione proteica dell’emoglobina può
modificare la capacità di legare ossigeno. Nell’uomo i cambiamenti fisiologici di pH, PCO2 e
temperatura del plasma modulano l’affinità di legame dell’emoglobina per l’ossigeno che di
conseguenza modificano la curva di dissociazione. L’aumento di temperatura, PCO2 o una
riduzione del pH diminuiscono l’affinità delle molecole di emoglobina per l’ossigeno e
spostano la curva verso destra. Dal punto di vista fisiologico, il livello di ossigeno a livello
polmonare non varia molto, mentre l’apporto di ossigeno a livello tissutale può essere
significativamente alterato.
ALTERAZIONE PH
Le alterazioni di pH nell’organismo avvengono per esempio in una situazione in cui l’esercizio
fisico al massimo dello sforzo spinge le cellule in anaerobiosi. Il metabolismo anaerobico
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
produce acido lattico, che rilascia H+. Quando il pH scende a causa degli ioni H+ l’affinità
dell’emoglobina all’ossigeno diminuisce e la curva di saturazione si sposta a destra. Quindi
viene rilasciata una maggiore quantità di ossigeno ai tessuti quando il sangue diventa più
acido e questo cambiamento è detto effetto Bohr.
ESEMPIO EMOGLOBINA FETALE
L’emoglobina fetale è costituita da due catene alfa e due gamma e questo appunto modifica
la struttura rendendola più affine all’O2, così nell’ambiente placentare l’O2 si stacca
dall’emoglobina materna ed è subito catturata dall’emoglobina fetale.
53. CONTROLLO RIFLESSO DELLA VENTILAZIONE DA PARTE DEI CHEMOCETTORI (PERIFERICI E
CENTRALI) E DEL CENTRO DI CONTROLLO DELLA RESPIRAZIONE BULBARE
La ventilazione è un processo ritmico che avviene involontariamente, ma può essere
controllata, fino ad un certo punto anche involontariamente.
Le informazioni sensoriali proveniente dai chemocettori centrali e periferici modificano la
ritmicità del generatore centrale di ritmi ventilatori contribuendo a mantenere l’omeostasi
dei gas nel sangue. L’anidride carbonica è lo stimolo principale che provoca modificazioni
della ventilazione, mentre l’ossigeno e il pH plasmatico svolgono un ruolo minore. I
chemocettori per l’ossigeno e per l’anidride carbonica sono associati alla circolazione
arteriosa. Se nel sangue arterioso è presente un livello di ossigeno troppo basso, la frequenza
e la profondità della ventilazione aumentano. Se la velocità di produzione dell’anidride
carbonica supera quella di rimozione polmonare, la PCO2 arteriosa aumenta e la ventilazione
è intensificata per equilibrare la rimozione di CO2 con la sua produzione. Questi riflessi
omeostatici operano costantemente, mantenendo la PO2 e la PCO2 arteriose all’interno di un
ambito ristretto di valori.
CHEMOCETTORI PERIFERICI: localizzati nelle arterie carotidi e aorta rilevano i cambiamenti di
PO2, pH e PCO2 nel plasma. Quando le cellule sono attivate da una diminuzione di PO2 o pH, o
da un aumento di PCO2, esse evocano un aumento riflesso della ventilazione. Una qualsiasi
condizione che determini una riduzione del pH plasmatico o un aumento della PCO2 attiverà
le cellule dei glomi carotidei e aortici e provocherà un aumento della ventilazione.
Chemocettori periferici rispondono solo a cambiamenti molto forti di PO2 arteriosa, essi non
hanno alcun ruolo nella regolazione quotidiana della ventilazione.
CHEMOCETTORI CENTRALI: la sostanza chimica più importante nel controllo della
ventilazione è l’anidride carbonica che agisce mediante i chemocettori periferici e i
chemocettori centrali situati nel bulbo. Questi recettori stabiliscono la frequenza ventilatoria,
fornendo continue informazioni al generatore centrale di ritmi ventilatori. Quando la PCO2
arteriosa aumenta, l’anidride carbonica attraversa la barriera emato – encefalica piuttosto
velocemente, provocando l’attivazione dei chemocettori centrali. Quest’ultimi segnalano al
generatore centrale di aumentare la frequenza e la profondità della ventilazione,
aumentando la ventilazione alveolare e rimuovendo l’anidride carbonica dal sangue. I
chemocettori centrali rispondono sia ad una riduzione della PCO2 che ad un suo aumento, se
la PCO2 alveolare diminuisce, la PCO2 nel plasma e quella nel liquido cerebrospinale
diminuiscono.
Document shared on www.docsity.com
Downloaded by: elia-cichetti (elia190cichetti@gmail.com)
Potrebbero piacerti anche
- Potenziale D'azioneDocumento30 paginePotenziale D'azioneRiccardo Mason TroiaNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- Guida pratica per ridurre le onde elettromagnetiche: Proteggersi dall’elettrosmog: tutti i rischi per la salute e come difendersiDa EverandGuida pratica per ridurre le onde elettromagnetiche: Proteggersi dall’elettrosmog: tutti i rischi per la salute e come difendersiNessuna valutazione finora
- Fisiopatologia Del Segnale Lezione 1 e 2Documento67 pagineFisiopatologia Del Segnale Lezione 1 e 2Mariagiovanna MariagiovannaNessuna valutazione finora
- FISIOLOGIAsbobineDocumento164 pagineFISIOLOGIAsbobinemarco.ricco.mr00Nessuna valutazione finora
- Il Potenziale D'azioneDocumento6 pagineIl Potenziale D'azioneLeandra EvolaNessuna valutazione finora
- Potenziali d'AzioneDocumento5 paginePotenziali d'AzioneHghh NgthhNessuna valutazione finora
- Il Sistema NervosoDocumento6 pagineIl Sistema NervosoMartina MusmeciNessuna valutazione finora
- Citologia 14 - Potenziale Di MembranaDocumento5 pagineCitologia 14 - Potenziale Di MembranaFrancesco TarulloNessuna valutazione finora
- Anche Ad Altre ComponentiDocumento4 pagineAnche Ad Altre Componentifrancesco biancoNessuna valutazione finora
- Appunti Fisiologia IDocumento27 pagineAppunti Fisiologia Ikekko88Nessuna valutazione finora
- Conduzione NervosaDocumento10 pagineConduzione NervosaMiriam LongobardiNessuna valutazione finora
- NEUROBIOLOGIA La SapienzaDocumento30 pagineNEUROBIOLOGIA La Sapienzachiara cimeiNessuna valutazione finora
- Fisiologia Lezione 4Documento6 pagineFisiologia Lezione 4The last chimeraNessuna valutazione finora
- Il Sistema NervosoDocumento8 pagineIl Sistema NervosoMartina RossiNessuna valutazione finora
- Conduzione Nervosa (Riassunto)Documento10 pagineConduzione Nervosa (Riassunto)Aaron RomanoNessuna valutazione finora
- 01-La Genesi Dell'impulso Nervoso PDFDocumento8 pagine01-La Genesi Dell'impulso Nervoso PDFDiego Flores EgüezNessuna valutazione finora
- Conduzione NervosaDocumento1 paginaConduzione NervosaeurusneNessuna valutazione finora
- FISIOLOGIADocumento30 pagineFISIOLOGIAlorenzo carliniNessuna valutazione finora
- FISIOLOGÍADocumento26 pagineFISIOLOGÍAAndrea De La FuenteNessuna valutazione finora
- Appunti Di NeurofisiologiaDocumento91 pagineAppunti Di NeurofisiologiaSaraNessuna valutazione finora
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- FisiologiaDocumento42 pagineFisiologiaLaura TreggiariNessuna valutazione finora
- Canali Ionici, Fibre NervoseDocumento22 pagineCanali Ionici, Fibre Nervoseapi-3704348100% (1)
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- Fisiologia Lezione 3Documento4 pagineFisiologia Lezione 3The last chimeraNessuna valutazione finora
- Sistema NervosoDocumento15 pagineSistema NervosoBruce WiiNessuna valutazione finora
- Segnali BioelettriciDocumento12 pagineSegnali BioelettriciFrancesco BorealeNessuna valutazione finora
- Potenziale Di MembranaDocumento17 paginePotenziale Di MembranamelissaNessuna valutazione finora
- Riassuntidi BiofisicaDocumento14 pagineRiassuntidi BiofisicaFlavia IovinoNessuna valutazione finora
- La Pompa Sodio-PotassioDocumento19 pagineLa Pompa Sodio-PotassioDavide ValmorbidaNessuna valutazione finora
- 1 Potenziale Di Equilibrio Di NernstDocumento5 pagine1 Potenziale Di Equilibrio Di NernstClaudia OrlannoNessuna valutazione finora
- Lezione 06-FisiologiaDocumento3 pagineLezione 06-FisiologiaSarah ScuderiNessuna valutazione finora
- Fisiologia II-7 - Il Sistema Di ConduzioneDocumento14 pagineFisiologia II-7 - Il Sistema Di ConduzioneShaun Ivan MuzicNessuna valutazione finora
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- Fisiologia Cellulare Potenziale AzioneDocumento31 pagineFisiologia Cellulare Potenziale Azionesimo_pNessuna valutazione finora
- Canali Ionici, Enzimi, Molecole CarrierDocumento114 pagineCanali Ionici, Enzimi, Molecole Carrierapi-26155364Nessuna valutazione finora
- Dispensa Fisiologia UnitoDocumento497 pagineDispensa Fisiologia Unitodorian88Nessuna valutazione finora
- CITOLOGIA E ISTOLOGIA - Lezione 7Documento2 pagineCITOLOGIA E ISTOLOGIA - Lezione 7Federico TosiniNessuna valutazione finora
- Potenziale D'azioneDocumento8 paginePotenziale D'azioneMicheleNessuna valutazione finora
- I Recettori SensorialiDocumento37 pagineI Recettori SensorialiMaryna100% (1)
- Sbobine Fisiologia Umana I 2015Documento443 pagineSbobine Fisiologia Umana I 2015Leonardo MarchettiNessuna valutazione finora
- Domande PIT FisiologiaDocumento5 pagineDomande PIT FisiologiaJoshuaKKyleNessuna valutazione finora
- 1 Potenziale Di MembranaDocumento2 pagine1 Potenziale Di MembranaGiacomo Ascari-RaccagniNessuna valutazione finora
- 5 Potenziali ElettrotoniciDocumento8 pagine5 Potenziali ElettrotoniciClaudia OrlannoNessuna valutazione finora
- Attività Elettrica Del CuoreDocumento26 pagineAttività Elettrica Del CuoreStella FilanteNessuna valutazione finora
- NeurofisiologiaDocumento69 pagineNeurofisiologiafabiana900% (1)
- Legge Di GHK, Comparazione Membrana - Condensatore, Reobase e Cronassia, Canali, Canali Pre Il SodioDocumento33 pagineLegge Di GHK, Comparazione Membrana - Condensatore, Reobase e Cronassia, Canali, Canali Pre Il Sodioapi-3704348100% (2)
- Bio Chi MicaDocumento59 pagineBio Chi Micafocaxo5386Nessuna valutazione finora
- FISIOLOGIA 1 Lezione UniDocumento6 pagineFISIOLOGIA 1 Lezione UniMariateresa SantorsolaNessuna valutazione finora
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- 02 Potenziale D'azioneDocumento13 pagine02 Potenziale D'azioneaxelshippoNessuna valutazione finora
- Conduzione Impulso NervosoDocumento32 pagineConduzione Impulso NervosoMarcoNessuna valutazione finora
- 10 - Potenziali Graduati e Potenziale D-AzioneDocumento48 pagine10 - Potenziali Graduati e Potenziale D-AzioneMarcoNessuna valutazione finora
- Fisiologia - Riassunti Kandel "Principi Di Neuroscienze" Cap. 6-15Documento32 pagineFisiologia - Riassunti Kandel "Principi Di Neuroscienze" Cap. 6-15Matteo Paolucci100% (15)
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- Valitutti Soluzioni Esercizi 82074 c3Documento4 pagineValitutti Soluzioni Esercizi 82074 c3marco poloNessuna valutazione finora
- Protocollo 1Documento4 pagineProtocollo 1dglivioNessuna valutazione finora
- DP Catalogue PDFDocumento7 pagineDP Catalogue PDFAlienshowNessuna valutazione finora
- SavanaDocumento19 pagineSavanaSilvia Giaretta100% (1)
- Cechov-Racconti Umoristici TESTODocumento77 pagineCechov-Racconti Umoristici TESTOLivio Liotta0% (1)