Potrebbero piacerti anche
- Potenziale D'azioneDocumento30 paginePotenziale D'azioneRiccardo Mason TroiaNessuna valutazione finora
- Sinapsi e NeurotrasmettitoriDocumento45 pagineSinapsi e NeurotrasmettitoriNicola Laurora0% (1)
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
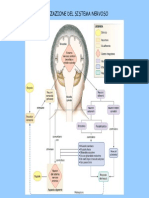
- Il Sistema NervosoDocumento6 pagineIl Sistema NervosoMartina MusmeciNessuna valutazione finora
- FISIOLOGIAsbobineDocumento164 pagineFISIOLOGIAsbobinemarco.ricco.mr00Nessuna valutazione finora
- 1 Potenziale Di Equilibrio Di NernstDocumento5 pagine1 Potenziale Di Equilibrio Di NernstClaudia OrlannoNessuna valutazione finora
- Fisiologia Cellulare Potenziale AzioneDocumento31 pagineFisiologia Cellulare Potenziale Azionesimo_pNessuna valutazione finora
- Docsity Domande Triennale Fisiologia UmanaDocumento33 pagineDocsity Domande Triennale Fisiologia Umanadestinyema78Nessuna valutazione finora
- 10 - Potenziali Graduati e Potenziale D-AzioneDocumento48 pagine10 - Potenziali Graduati e Potenziale D-AzioneMarcoNessuna valutazione finora
- Sistema NervosoDocumento15 pagineSistema NervosoBruce WiiNessuna valutazione finora
- L'impulso NervosoDocumento7 pagineL'impulso NervosoGiancarlo AttivissimoNessuna valutazione finora
- Potenziali D'azioneDocumento5 paginePotenziali D'azioneHghh NgthhNessuna valutazione finora
- Il Sistema NervosoDocumento8 pagineIl Sistema NervosoMartina RossiNessuna valutazione finora
- Appunti Di NeurofisiologiaDocumento91 pagineAppunti Di NeurofisiologiaSaraNessuna valutazione finora
- 4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azioneDocumento43 pagine4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azionedavide di lorenzoNessuna valutazione finora
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- 2.impulso NervosoDocumento10 pagine2.impulso NervosofNessuna valutazione finora
- CITOLOGIA E ISTOLOGIA - Lezione 7Documento2 pagineCITOLOGIA E ISTOLOGIA - Lezione 7Federico TosiniNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- 1 AprDocumento35 pagine1 Apralison.pedrini98Nessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- NEUROBIOLOGIA La SapienzaDocumento30 pagineNEUROBIOLOGIA La Sapienzachiara cimeiNessuna valutazione finora
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- Conduzione Nervosa (Riassunto)Documento10 pagineConduzione Nervosa (Riassunto)Aaron RomanoNessuna valutazione finora
- 01-La Genesi Dell'impulso Nervoso PDFDocumento8 pagine01-La Genesi Dell'impulso Nervoso PDFDiego Flores EgüezNessuna valutazione finora
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- Lezione 2 FisiologiaDocumento47 pagineLezione 2 Fisiologiamarticatalin8Nessuna valutazione finora
- La Pompa Sodio-PotassioDocumento19 pagineLa Pompa Sodio-PotassioDavide ValmorbidaNessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- Conduzione Impulso NervosoDocumento32 pagineConduzione Impulso NervosoMarcoNessuna valutazione finora
- Potenziale AzioneDocumento30 paginePotenziale AzionePatrizio SpadoniNessuna valutazione finora
- Il Potenziale D'azioneDocumento6 pagineIl Potenziale D'azioneLeandra EvolaNessuna valutazione finora
- Lezione 06-FisiologiaDocumento3 pagineLezione 06-FisiologiaSarah ScuderiNessuna valutazione finora
- Sbobine Fisiologia Umana I 2015Documento443 pagineSbobine Fisiologia Umana I 2015Leonardo MarchettiNessuna valutazione finora
- Conduzione NervosaDocumento1 paginaConduzione NervosaeurusneNessuna valutazione finora
- Dispensa Fisiologia UnitoDocumento497 pagineDispensa Fisiologia Unitodorian88Nessuna valutazione finora
- I Recettori SensorialiDocumento37 pagineI Recettori SensorialiMaryna100% (1)
- FisiologiaDocumento42 pagineFisiologiaLaura TreggiariNessuna valutazione finora
- Potenziale Di MembranaDocumento17 paginePotenziale Di MembranamelissaNessuna valutazione finora
- Trasmissione Sinaptica e Integrazione NeuronaleDocumento11 pagineTrasmissione Sinaptica e Integrazione NeuronaleAnnamaria MaderaNessuna valutazione finora
- Anche Ad Altre ComponentiDocumento4 pagineAnche Ad Altre Componentifrancesco biancoNessuna valutazione finora
- Occhio3 Recettori Corteccia VieDocumento43 pagineOcchio3 Recettori Corteccia Viesimo_pNessuna valutazione finora
- Segnali BioelettriciDocumento12 pagineSegnali BioelettriciFrancesco BorealeNessuna valutazione finora
- EvoliDocumento93 pagineEvoliPietro BracagliaNessuna valutazione finora
- Domande PIT FisiologiaDocumento5 pagineDomande PIT FisiologiaJoshuaKKyleNessuna valutazione finora
- Citologia 14 - Potenziale Di MembranaDocumento5 pagineCitologia 14 - Potenziale Di MembranaFrancesco TarulloNessuna valutazione finora
- NeurofisiologiaDocumento69 pagineNeurofisiologiafabiana900% (1)
- 2 - BioelettricitàDocumento4 pagine2 - BioelettricitàFabio BordinNessuna valutazione finora
- Fisiologia ValenteDocumento4 pagineFisiologia ValenteFrancesco FalconieriNessuna valutazione finora
- Fisiologia Dei Sistemi MotoriDocumento17 pagineFisiologia Dei Sistemi Motorisara.zandonai99Nessuna valutazione finora
- Tessuto NervosoDocumento7 pagineTessuto NervosoeristelaNessuna valutazione finora
- Fisiologia SinapsiDocumento1 paginaFisiologia SinapsiFedericaRomanoNessuna valutazione finora
- Fisiologia PDFDocumento75 pagineFisiologia PDFMarioNessuna valutazione finora
- Intro Eeg BciDocumento28 pagineIntro Eeg BciHibino TsubakiNessuna valutazione finora
- Riassuntidi BiofisicaDocumento14 pagineRiassuntidi BiofisicaFlavia IovinoNessuna valutazione finora
- Canali Ionici, Enzimi, Molecole CarrierDocumento114 pagineCanali Ionici, Enzimi, Molecole Carrierapi-26155364Nessuna valutazione finora
- 5 Potenziali ElettrotoniciDocumento8 pagine5 Potenziali ElettrotoniciClaudia OrlannoNessuna valutazione finora
- FISIOLOGIADocumento30 pagineFISIOLOGIAlorenzo carliniNessuna valutazione finora
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- Nanotecnologie per il trattamento dei tumori: quali sono gli effetti sul genoma?Da EverandNanotecnologie per il trattamento dei tumori: quali sono gli effetti sul genoma?Nessuna valutazione finora