Potrebbero piacerti anche
- Interpretazione dell'ECG: Una Guida Pratica ed Intuitiva per Imparare a Leggere l'ECG e a Diagnosticare e Trattare le AritmieDa EverandInterpretazione dell'ECG: Una Guida Pratica ed Intuitiva per Imparare a Leggere l'ECG e a Diagnosticare e Trattare le AritmieNessuna valutazione finora
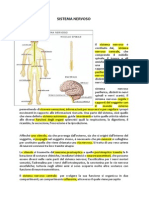
- Scienze - Il Sistema NervosoDocumento27 pagineScienze - Il Sistema NervosoRiccardo PengoNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- 120 Domande FisiologiaDocumento6 pagine120 Domande FisiologiagoyshorvalNessuna valutazione finora
- Muscolo LiscioDocumento14 pagineMuscolo LiscioDaniela PiteoNessuna valutazione finora
- UntitledDocumento14 pagineUntitledMartina ZuccariniNessuna valutazione finora
- Canali Ionici, Fibre NervoseDocumento22 pagineCanali Ionici, Fibre Nervoseapi-3704348100% (1)
- Messaggeri ChimiciDocumento30 pagineMessaggeri ChimiciMarynaNessuna valutazione finora
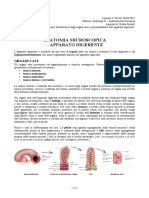
- 18 - Anatomia II (Microscopica) - 13.03.2017 - RDocumento8 pagine18 - Anatomia II (Microscopica) - 13.03.2017 - RSandroNessuna valutazione finora
- NeurotrasmettitoriDocumento4 pagineNeurotrasmettitoriMarco ParolinNessuna valutazione finora
- Appunti Fisiologia IDocumento27 pagineAppunti Fisiologia Ikekko88Nessuna valutazione finora
- 4 Lezione - La Cute e I Suoi AnnessiDocumento24 pagine4 Lezione - La Cute e I Suoi AnnessiAlbert stefan PaulencoNessuna valutazione finora
- Fisiologia AppuntiDocumento20 pagineFisiologia AppuntienzocavajuoloNessuna valutazione finora
- Appunti FisiologiaDocumento83 pagineAppunti FisiologiaWalter ArmeliNessuna valutazione finora
- Biochimica GeneraleDocumento29 pagineBiochimica Generalenikkamclean100% (1)
- Sistema EndocrinoDocumento34 pagineSistema EndocrinoDiego LorussoNessuna valutazione finora
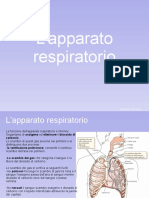
- Il Sistema RespiratorioDocumento15 pagineIl Sistema RespiratorioCaterina RizzoNessuna valutazione finora
- (Med - Ita - Istologia Sintesi - EmbriologiaDocumento31 pagine(Med - Ita - Istologia Sintesi - EmbriologiammNessuna valutazione finora
- Svolti 3Documento91 pagineSvolti 3DeejayLupinNessuna valutazione finora
- Gas Respiratori, Rapporto Tra Ventilazione e PerfusioneDocumento15 pagineGas Respiratori, Rapporto Tra Ventilazione e Perfusioneapi-37043480% (1)
- Apparato UrinarioDocumento17 pagineApparato UrinarioCate?Nessuna valutazione finora
- 26 - Anatomia II - 30-03-2017 (R)Documento10 pagine26 - Anatomia II - 30-03-2017 (R)SandroNessuna valutazione finora
- Elenco Quesiti Biochimica ClinicaDocumento10 pagineElenco Quesiti Biochimica ClinicaNicola LauroraNessuna valutazione finora
- Lezione II-10 Endocrino 04-04-11Documento91 pagineLezione II-10 Endocrino 04-04-11Roberta RossiNessuna valutazione finora
- 30 Anatomia II 02.03.2016 Laringe e MediastinoDocumento7 pagine30 Anatomia II 02.03.2016 Laringe e MediastinoSandroNessuna valutazione finora
- Introduzione TEM 2021Documento15 pagineIntroduzione TEM 2021TomyasuNessuna valutazione finora
- 1163 - La Membrana Plasmatica PDFDocumento8 pagine1163 - La Membrana Plasmatica PDFDiego Flores EgüezNessuna valutazione finora
- Mappa Concettuale Sull'Apparato RespiratorioDocumento1 paginaMappa Concettuale Sull'Apparato RespiratorioBOLFRA100% (1)
- Sistema Vestibolare, Funzioni Superiori Dell'encefalo, LinguaggioDocumento17 pagineSistema Vestibolare, Funzioni Superiori Dell'encefalo, Linguaggioapi-3704348Nessuna valutazione finora
- 31-Anatomia II-03.03.2016-Topografia Anatomica Del CuoreDocumento6 pagine31-Anatomia II-03.03.2016-Topografia Anatomica Del CuoreSandroNessuna valutazione finora
- Fisiologia Del Muscolo ScheletricoDocumento20 pagineFisiologia Del Muscolo ScheletricoCarlo BritoNessuna valutazione finora
- Sistema NervosoDocumento36 pagineSistema NervosoalinabortaNessuna valutazione finora
- Anatomia SNPDocumento30 pagineAnatomia SNPantonio nennaNessuna valutazione finora
- Enzimi PlasmaticiDocumento38 pagineEnzimi Plasmaticiapi-3704348100% (1)
- Appunti Sulla Duplicazione Del DNA: Trascrizione e TraduzioneDocumento6 pagineAppunti Sulla Duplicazione Del DNA: Trascrizione e TraduzionePiccolo GiuseppeNessuna valutazione finora
- 24 - Anatomia II - 27-03-2017Documento12 pagine24 - Anatomia II - 27-03-2017SandroNessuna valutazione finora
- Appunti MicrobiologiaDocumento79 pagineAppunti MicrobiologiaAmaiableachNessuna valutazione finora
- SNP Plesso Cervicale e BrachialeDocumento7 pagineSNP Plesso Cervicale e BrachialeMichelangelo Scanu100% (1)
- Apparato Respiratorio e SpirometriaDocumento9 pagineApparato Respiratorio e Spirometriaapi-3704348100% (1)
- Flusso Respiratorio, O2 e CO2Documento9 pagineFlusso Respiratorio, O2 e CO2api-3704348100% (3)
- 30 - Anatomia II - 26-04-2017 - RDocumento11 pagine30 - Anatomia II - 26-04-2017 - RSandroNessuna valutazione finora
- Sbobina Chimica (1) - 1Documento239 pagineSbobina Chimica (1) - 1Roberta VisoneNessuna valutazione finora
- 36-Anatomia II-16.03.16-Anatomia Strutturistica e MicroscopicaDocumento6 pagine36-Anatomia II-16.03.16-Anatomia Strutturistica e MicroscopicaSandroNessuna valutazione finora
- Il Sistema Nervoso SensorialeDocumento69 pagineIl Sistema Nervoso Sensorialesimo_p100% (1)
- 13 - Immunologia-26.04.2017Documento11 pagine13 - Immunologia-26.04.2017SandroNessuna valutazione finora
- Lezioni Vie Nervose SNAutonomoDocumento80 pagineLezioni Vie Nervose SNAutonomoVincenzo BinantiNessuna valutazione finora
- Massimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare PolmonareDocumento21 pagineMassimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare Polmonareapi-3704348100% (1)
- Le Ghiandole IIDocumento20 pagineLe Ghiandole IIEleonora Di MarcoNessuna valutazione finora
- Apparato RespiratorioDocumento9 pagineApparato Respiratoriochiara9rossoNessuna valutazione finora
- Lo Stress - Tesi Di BarbaraDocumento46 pagineLo Stress - Tesi Di BarbaraSalvatore BonaffinoNessuna valutazione finora
- Fisiologia GeneraleDocumento158 pagineFisiologia GeneraleSusannaNessuna valutazione finora
- Immunologia ClinicaDocumento38 pagineImmunologia ClinicaFedericoPppzPippo100% (1)
- L'Asse Ipotalamo - Ipofisario e L'epifisiDocumento12 pagineL'Asse Ipotalamo - Ipofisario e L'epifisiGabriele Di SienaNessuna valutazione finora
- Apparato DigerenteDocumento47 pagineApparato Digerentelullaby02Nessuna valutazione finora
- Stimolo e Risposta Muscolare, RiflessoDocumento16 pagineStimolo e Risposta Muscolare, Riflessoapi-3704348100% (2)
- Neuroanatomia SchemiDocumento8 pagineNeuroanatomia SchemiHeroInLifeNessuna valutazione finora
- IL POTERE DELLA SEMPLICITA' Manuale di esercizi per l'autorealizzazioneDa EverandIL POTERE DELLA SEMPLICITA' Manuale di esercizi per l'autorealizzazioneNessuna valutazione finora
- Il trattamento della cefalea e dell'emicrania: ipotesi etiopatogenetiche ed efficacia dell'intervento osteopaticoDa EverandIl trattamento della cefalea e dell'emicrania: ipotesi etiopatogenetiche ed efficacia dell'intervento osteopaticoNessuna valutazione finora
- Risposte Savi. OdtDocumento2 pagineRisposte Savi. OdtNicola LauroraNessuna valutazione finora
- Chirurgia Pediatrica Lez 17Documento28 pagineChirurgia Pediatrica Lez 17Nicola LauroraNessuna valutazione finora
- Menu La Birretta Ascoli Di GiugnoDocumento9 pagineMenu La Birretta Ascoli Di GiugnoNicola LauroraNessuna valutazione finora
- Secondo InvitoDocumento1 paginaSecondo InvitoNicola LauroraNessuna valutazione finora
- 11 Etichette Birra Weizen LeviosaDocumento1 pagina11 Etichette Birra Weizen LeviosaNicola LauroraNessuna valutazione finora
- Gio V TimetableDocumento1 paginaGio V TimetableNicola LauroraNessuna valutazione finora
- DOMANDE GeneticaDocumento4 pagineDOMANDE GeneticaNicola LauroraNessuna valutazione finora
- Chir Vascolare PDFDocumento44 pagineChir Vascolare PDFNicola LauroraNessuna valutazione finora
- Guida Alla Fermentazione 14Documento12 pagineGuida Alla Fermentazione 14Nicola LauroraNessuna valutazione finora
- Principi Di FarmacocineticaDocumento36 paginePrincipi Di FarmacocineticaNicola LauroraNessuna valutazione finora
- Istruzioni Koji MR MaltDocumento3 pagineIstruzioni Koji MR MaltNicola LauroraNessuna valutazione finora
- MenecmiDocumento32 pagineMenecmidarkdeathmoonNessuna valutazione finora
- Med Ita - Metabolismo LipidicoDocumento3 pagineMed Ita - Metabolismo LipidicoNicola LauroraNessuna valutazione finora
- APPARATO DIGERENTE ChirurguaDocumento52 pagineAPPARATO DIGERENTE ChirurguaNicola LauroraNessuna valutazione finora
- Avviso Pagina in AggiornamentoDocumento1 paginaAvviso Pagina in AggiornamentoNicola LauroraNessuna valutazione finora
- Domande Chirurgia ToracicaDocumento45 pagineDomande Chirurgia ToracicaNicola LauroraNessuna valutazione finora
- Chirurgia GeneraleDocumento53 pagineChirurgia GeneraleNicola LauroraNessuna valutazione finora
- A Tutto RisottoDocumento21 pagineA Tutto RisottoVitina MaioNessuna valutazione finora
- Canti e Metodi Nel Ba Gua ZhangDocumento3 pagineCanti e Metodi Nel Ba Gua ZhangNicola LauroraNessuna valutazione finora
- Farmacologia TossicodipendenzeDocumento20 pagineFarmacologia TossicodipendenzeNicola LauroraNessuna valutazione finora
- Il BaguazhangDocumento2 pagineIl BaguazhangNicola LauroraNessuna valutazione finora
- Cucina Regionale ADocumento67 pagineCucina Regionale AAna BarrosNessuna valutazione finora
- Med Ita - Sistema NervosoDocumento40 pagineMed Ita - Sistema NervosoNicola LauroraNessuna valutazione finora
- Snami LegaleDocumento42 pagineSnami LegaleNicola LauroraNessuna valutazione finora
- Principi Di FarmacocineticaDocumento36 paginePrincipi Di FarmacocineticaNicola LauroraNessuna valutazione finora
- Med Ita - Metabolismo GlicidicoDocumento3 pagineMed Ita - Metabolismo GlicidicoNicola LauroraNessuna valutazione finora
- Principi Di FarmacocineticaDocumento36 paginePrincipi Di FarmacocineticaNicola LauroraNessuna valutazione finora
- Med Ita - GastrointestinaliDocumento6 pagineMed Ita - GastrointestinaliNicola LauroraNessuna valutazione finora
- Domande Medico-Infermiere-SanitarioDocumento3 pagineDomande Medico-Infermiere-SanitarioNicola LauroraNessuna valutazione finora
- Farmacologia TossicodipendenzeDocumento20 pagineFarmacologia TossicodipendenzeNicola LauroraNessuna valutazione finora
- Foglia IDocumento14 pagineFoglia IAnna De SimoneNessuna valutazione finora
- Analisi Citometriche NeoplasieDocumento295 pagineAnalisi Citometriche NeoplasielucioformigoniNessuna valutazione finora
- Malattie Autoimmuni: Storia Di Un Malinteso : Giancarlo CavallinoDocumento8 pagineMalattie Autoimmuni: Storia Di Un Malinteso : Giancarlo CavallinoNino BallarinoNessuna valutazione finora
- Neuroscienze CognitiveDocumento8 pagineNeuroscienze CognitiveLorenzo Maria CerviNessuna valutazione finora
- LA MICROIMMUNOTERAPIA LABO' LIFE Come Terapia Di Supporto Nelle Patologie Cronico - Degenerative e Nelle Patologie AutoimmuniDocumento259 pagineLA MICROIMMUNOTERAPIA LABO' LIFE Come Terapia Di Supporto Nelle Patologie Cronico - Degenerative e Nelle Patologie Autoimmuniweb3351Nessuna valutazione finora