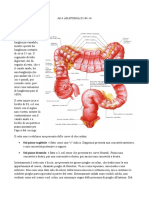
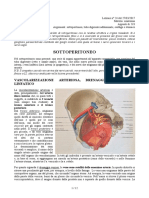
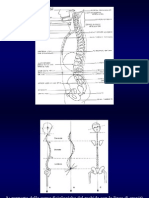
Potrebbero piacerti anche
- 1 - Sensibilità SomestesicaDocumento23 pagine1 - Sensibilità Somestesicaapi-3704348100% (3)
- Sistema NervosoDocumento26 pagineSistema NervosoTessy PsibarNessuna valutazione finora
- Propriocezione e Movimento (Appunti Del 2003, 14 Pagine) PDFDocumento14 paginePropriocezione e Movimento (Appunti Del 2003, 14 Pagine) PDFrosaNessuna valutazione finora
- Muscolo e LocomozioneDocumento21 pagineMuscolo e Locomozioneapi-3704348100% (1)
- Dispensa NeuroanatomiaDocumento142 pagineDispensa NeuroanatomiaPierpaolo LuzioNessuna valutazione finora
- Allenamento, fitness e bodybuilding: La guida definitiva per l’aumento della massa muscolare scegliendo schede e giusti esercizi da svolgere in palestra o a casa, con pesi, macchine o a corpo liberoDa EverandAllenamento, fitness e bodybuilding: La guida definitiva per l’aumento della massa muscolare scegliendo schede e giusti esercizi da svolgere in palestra o a casa, con pesi, macchine o a corpo liberoNessuna valutazione finora
- Gli Stupefacenti Segreti dello Yogi - (Tradotto)Da EverandGli Stupefacenti Segreti dello Yogi - (Tradotto)Nessuna valutazione finora
- Virilità: Come conservare e accrescere le forze virili - Cura dell'impotenza fisica e nervosaDa EverandVirilità: Come conservare e accrescere le forze virili - Cura dell'impotenza fisica e nervosaNessuna valutazione finora
- Cause, evoluzione e trattamento osteopatico delle distorsioni del rachide cervicaleDa EverandCause, evoluzione e trattamento osteopatico delle distorsioni del rachide cervicaleNessuna valutazione finora
- Diaframma, Rieducazione.Documento2 pagineDiaframma, Rieducazione.Tfames3274Nessuna valutazione finora
- Introduzione TEM 2021Documento15 pagineIntroduzione TEM 2021TomyasuNessuna valutazione finora
- GnatologiaDocumento38 pagineGnatologiaFrancesco SonnatiNessuna valutazione finora
- Metodo MezieresDocumento33 pagineMetodo MezieresRoberto DoreNessuna valutazione finora
- Introduzione Fasce 2021Documento32 pagineIntroduzione Fasce 2021TomyasuNessuna valutazione finora
- Sistema Muscolo-ScheletricoDocumento6 pagineSistema Muscolo-ScheletricoturchettinoNessuna valutazione finora
- Rubrica-Osteopatia 1 CostaDocumento2 pagineRubrica-Osteopatia 1 Costapaolo68audisioNessuna valutazione finora
- Sindrome Dello Stretto Toracico SuperioreDocumento5 pagineSindrome Dello Stretto Toracico SuperioremaxNessuna valutazione finora
- Curare I Dolori Con La Riflessologia PlantareDocumento16 pagineCurare I Dolori Con La Riflessologia PlantareclekupreNessuna valutazione finora
- Tesi SplattoDocumento42 pagineTesi SplattoGiuseppe02Nessuna valutazione finora
- SNP Plesso Cervicale e BrachialeDocumento7 pagineSNP Plesso Cervicale e BrachialeMichelangelo Scanu100% (1)
- 16 Anatomia 02-04-14Documento8 pagine16 Anatomia 02-04-14SandroNessuna valutazione finora
- Anatomia 2 (Denise) AppuntiunipiDocumento158 pagineAnatomia 2 (Denise) AppuntiunipinuunumclNessuna valutazione finora
- 24 - Anatomia II - 27-03-2017Documento12 pagine24 - Anatomia II - 27-03-2017SandroNessuna valutazione finora
- Nervi Cranici FinaleDocumento30 pagineNervi Cranici FinalefedericoNessuna valutazione finora
- Agopuntura Nella Patologia Della SpallaDocumento4 pagineAgopuntura Nella Patologia Della SpallaAnnunzio MatráNessuna valutazione finora
- 1413140517disturbi e Osteopatia PDFDocumento12 pagine1413140517disturbi e Osteopatia PDFpaolo68audisioNessuna valutazione finora
- 6 - Controllo MotorioDocumento17 pagine6 - Controllo Motorioapi-3704348100% (2)
- Terapia Fisica Della Spalla (Donatelli)Documento444 pagineTerapia Fisica Della Spalla (Donatelli)AlessandroNessuna valutazione finora
- Rieducazione SpallaDocumento7 pagineRieducazione SpallateresaioanaNessuna valutazione finora
- Fisiopatologia Del Sistema MotorioDocumento58 pagineFisiopatologia Del Sistema MotorioIlaria PuttiniNessuna valutazione finora
- Viscerale Lezione 1 Parte 2Documento13 pagineViscerale Lezione 1 Parte 2Antonio GarNessuna valutazione finora
- Fascia CervicaleDocumento14 pagineFascia CervicaleFrancesco Valsega100% (1)
- Massaggio SportivoDocumento2 pagineMassaggio Sportivolauretta100Nessuna valutazione finora
- Plessi EOMDocumento7 paginePlessi EOMPery1945Nessuna valutazione finora
- Lezioni Vie Nervose SNAutonomoDocumento80 pagineLezioni Vie Nervose SNAutonomoVincenzo BinantiNessuna valutazione finora
- 43-Anatomia II-14.04.2016 - Fegato, Pancreas e MilzaDocumento7 pagine43-Anatomia II-14.04.2016 - Fegato, Pancreas e MilzaSandroNessuna valutazione finora
- Il Sovraccarico Del GinocchioDocumento8 pagineIl Sovraccarico Del GinocchiocactusmtbNessuna valutazione finora
- 1 Lezione - I Tessuti EpitelialiDocumento16 pagine1 Lezione - I Tessuti EpitelialiAlbert stefan PaulencoNessuna valutazione finora
- Rachide LombareDocumento4 pagineRachide Lombareelpibedeoro1993Nessuna valutazione finora
- La Potenza Della QuieteDocumento28 pagineLa Potenza Della QuieteDavide MagliarisiNessuna valutazione finora
- Stress Benessere Giovanni ChettaDocumento27 pagineStress Benessere Giovanni ChettaD. S.Nessuna valutazione finora
- Stimolo e Risposta Muscolare, RiflessoDocumento16 pagineStimolo e Risposta Muscolare, Riflessoapi-3704348100% (2)
- 36-Anatomia II-16.03.16-Anatomia Strutturistica e MicroscopicaDocumento6 pagine36-Anatomia II-16.03.16-Anatomia Strutturistica e MicroscopicaSandroNessuna valutazione finora
- Varie Patologie Del Pavimento PelvicoDocumento37 pagineVarie Patologie Del Pavimento PelvicoDaniel TacheNessuna valutazione finora
- Corso Integrato Istologia e Anatomia Umana I Per MedicinaDocumento6 pagineCorso Integrato Istologia e Anatomia Umana I Per MedicinaGiamma SinisiNessuna valutazione finora
- Biomeccanica Anatomo FunzionaleDocumento257 pagineBiomeccanica Anatomo FunzionaleFlavio VillaNessuna valutazione finora
- Stretching - Metodologie Funzionali e Loro ApplicazioneDocumento11 pagineStretching - Metodologie Funzionali e Loro ApplicazioneAlessio FerlitoNessuna valutazione finora
- Delle Manipolazioni Vertebrali Con La Tensegrita by Salmochi Jean FrançoisDocumento2 pagineDelle Manipolazioni Vertebrali Con La Tensegrita by Salmochi Jean FrançoisTensegrity WikiNessuna valutazione finora
- Massimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare PolmonareDocumento21 pagineMassimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare Polmonareapi-3704348100% (1)
- 2 Lezione - I Tessuti ConnettiviDocumento19 pagine2 Lezione - I Tessuti ConnettiviAlbert stefan PaulencoNessuna valutazione finora
- Acupuntura e Book Ita Agopuntura Elenco Dei PuntiDocumento20 pagineAcupuntura e Book Ita Agopuntura Elenco Dei PuntiAurea813Nessuna valutazione finora
- Imaging RachideDocumento59 pagineImaging Rachidethe_walnutNessuna valutazione finora
- Viscerale Lezione 1 Parte 3Documento9 pagineViscerale Lezione 1 Parte 3Antonio GarNessuna valutazione finora
- DellaFisonomiadellHuomo 11162694 PDFDocumento503 pagineDellaFisonomiadellHuomo 11162694 PDFChukno ChuknoNessuna valutazione finora
- 20 - Anatomia II - 16.03.2017 - RDocumento10 pagine20 - Anatomia II - 16.03.2017 - RSandroNessuna valutazione finora
- 4 Lezione - La Cute e I Suoi AnnessiDocumento24 pagine4 Lezione - La Cute e I Suoi AnnessiAlbert stefan PaulencoNessuna valutazione finora
- Esercizi Metodo MckenzieDocumento5 pagineEsercizi Metodo Mckenzie12345678hjNessuna valutazione finora
- Appunti Di Ortopedia e TraumatologiaDocumento43 pagineAppunti Di Ortopedia e TraumatologiaLuciano BecciaNessuna valutazione finora
- Schema CorporeoDocumento10 pagineSchema CorporeoAlessandroVolpe100% (1)
- Controllo Dell'attività Cardiaca e RespiratoriaDocumento22 pagineControllo Dell'attività Cardiaca e Respiratoriaapi-3704348100% (4)
- Circolo Polmonare, Effetto Waterfall, Legge Di Starling, Sistema LinfaticoDocumento16 pagineCircolo Polmonare, Effetto Waterfall, Legge Di Starling, Sistema Linfaticoapi-3704348Nessuna valutazione finora
- Test 4Documento10 pagineTest 4api-3704348100% (1)
- Test 1Documento8 pagineTest 1api-3704348Nessuna valutazione finora
- Cuore, Arteriole, Distretto Venoso, ECGDocumento13 pagineCuore, Arteriole, Distretto Venoso, ECGapi-3704348100% (1)
- Es5 QuizDocumento12 pagineEs5 Quizapi-3704348Nessuna valutazione finora
- Epiteli SensorialiDocumento5 pagineEpiteli Sensorialiapi-3704348100% (1)
- Caratteristiche Del MuscoloDocumento16 pagineCaratteristiche Del Muscoloapi-3704348100% (3)
- Calcolo Dello Shunt, Consumo Di Ossigenocontrollo Nervoso e Chimico Della RespirazioneDocumento23 pagineCalcolo Dello Shunt, Consumo Di Ossigenocontrollo Nervoso e Chimico Della Respirazioneapi-3704348100% (5)
- Test 5 (Trascritto A Mano)Documento13 pagineTest 5 (Trascritto A Mano)api-3704348Nessuna valutazione finora
- Test 2Documento7 pagineTest 2api-3704348Nessuna valutazione finora
- Test 3Documento8 pagineTest 3api-3704348Nessuna valutazione finora
- Es4 QuizDocumento3 pagineEs4 Quizapi-3704348Nessuna valutazione finora
- Test 6 (Trascritto A Mano)Documento8 pagineTest 6 (Trascritto A Mano)api-3704348Nessuna valutazione finora
- Es1 QuizDocumento4 pagineEs1 Quizapi-3704348Nessuna valutazione finora
- Es2 QuizDocumento11 pagineEs2 Quizapi-3704348Nessuna valutazione finora
- Es6 QuizDocumento10 pagineEs6 Quizapi-3704348Nessuna valutazione finora
- Es3 QuizDocumento11 pagineEs3 Quizapi-3704348Nessuna valutazione finora
- Neurofisio - Sinapsi, Secondi Messaggeri NeurotrasmettitoriDocumento24 pagineNeurofisio - Sinapsi, Secondi Messaggeri Neurotrasmettitoriapi-3704348100% (2)
- Sinapsi Recettori Leggi Di Weber e Stevens Tatto e Corpuscoli Recettoriali Inibizione LateraleBNDocumento20 pagineSinapsi Recettori Leggi Di Weber e Stevens Tatto e Corpuscoli Recettoriali Inibizione LateraleBNLaura MedinaNessuna valutazione finora
- Via Ottica e Fisiologia Dei Recettori Visivi - BNDocumento36 pagineVia Ottica e Fisiologia Dei Recettori Visivi - BNapi-3704348100% (1)
- Ottica Seconda ParteDocumento24 pagineOttica Seconda Parteapi-3704348Nessuna valutazione finora
- Il Lab Oratorio Di Sierologia e Di VirologiaDocumento11 pagineIl Lab Oratorio Di Sierologia e Di Virologiaapi-3704348Nessuna valutazione finora
- Gusto e Olfatto - BNDocumento7 pagineGusto e Olfatto - BNapi-3704348Nessuna valutazione finora
- Compito GiugnoDocumento5 pagineCompito Giugnoapi-3704348100% (1)
- AntibodiesDocumento8 pagineAntibodiesapi-3704348Nessuna valutazione finora
- Canali Ionici, Fibre Nervose, Velocità Di Conduzione - BNDocumento29 pagineCanali Ionici, Fibre Nervose, Velocità Di Conduzione - BNapi-3704348Nessuna valutazione finora
- Prove Esame Immuno 1Documento19 pagineProve Esame Immuno 1api-3704348Nessuna valutazione finora
- Programma Con ImmaginiDocumento2 pagineProgramma Con Immaginiapi-3704348Nessuna valutazione finora
- Immuno DiagnosiDocumento13 pagineImmuno Diagnosiapi-3704348Nessuna valutazione finora