Potrebbero piacerti anche
- Il Tuina: La più antica arte manipolativa dei tessuti biologici per il raggiungimento dello stato di buona salute psico-fisicaDa EverandIl Tuina: La più antica arte manipolativa dei tessuti biologici per il raggiungimento dello stato di buona salute psico-fisicaNessuna valutazione finora
- TesiDocumento62 pagineTesiSimonaNessuna valutazione finora
- Allenamento, fitness e bodybuilding: La guida definitiva per l’aumento della massa muscolare scegliendo schede e giusti esercizi da svolgere in palestra o a casa, con pesi, macchine o a corpo liberoDa EverandAllenamento, fitness e bodybuilding: La guida definitiva per l’aumento della massa muscolare scegliendo schede e giusti esercizi da svolgere in palestra o a casa, con pesi, macchine o a corpo liberoNessuna valutazione finora
- Lo Stress - Tesi Di BarbaraDocumento46 pagineLo Stress - Tesi Di BarbaraSalvatore BonaffinoNessuna valutazione finora
- Le Ghiandole IIDocumento20 pagineLe Ghiandole IIEleonora Di MarcoNessuna valutazione finora
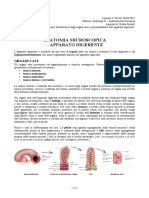
- Apparato DigerenteDocumento47 pagineApparato Digerentelullaby02Nessuna valutazione finora
- La Fisiopatologia Muscolare - DefDocumento28 pagineLa Fisiopatologia Muscolare - Defgiuliacagnina1Nessuna valutazione finora
- Respirazione Addominale AppuntiDocumento4 pagineRespirazione Addominale AppuntiLaerteNessuna valutazione finora
- La Circolazione SanguignaDocumento4 pagineLa Circolazione SanguignaManuela MarraNessuna valutazione finora
- Protocollo Riabilitativo Anca (DA FARE)Documento6 pagineProtocollo Riabilitativo Anca (DA FARE)Flavia IovinoNessuna valutazione finora
- Il Sistema RespiratorioDocumento12 pagineIl Sistema RespiratorioFabiana FonsinoNessuna valutazione finora
- Cap Mammella X27592allp1Documento10 pagineCap Mammella X27592allp1Raluca BordeianuNessuna valutazione finora
- La Persona in Psicologia Sociale - Burr PDFDocumento58 pagineLa Persona in Psicologia Sociale - Burr PDFIgnacio VenturiniNessuna valutazione finora
- Panoramica Sulle Distorsioni e Altre Lesioni Dei Tessuti Molli - Lesioni e Avvelenamento - Manuale MSD, Versione Per I PazientiDocumento13 paginePanoramica Sulle Distorsioni e Altre Lesioni Dei Tessuti Molli - Lesioni e Avvelenamento - Manuale MSD, Versione Per I PazientiBeppe FranchinoNessuna valutazione finora
- Aerobica TraumatologiaDocumento14 pagineAerobica TraumatologiacactusmtbNessuna valutazione finora
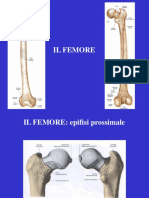
- 04.3 Lo Scheletro e Le Articolazioni Dell Arto InferioreDocumento61 pagine04.3 Lo Scheletro e Le Articolazioni Dell Arto InferioreAlucardNessuna valutazione finora
- Fisiologia Del Muscolo ScheletricoDocumento20 pagineFisiologia Del Muscolo ScheletricoCarlo BritoNessuna valutazione finora
- Muscolo LiscioDocumento14 pagineMuscolo LiscioDaniela PiteoNessuna valutazione finora
- 30 - Anatomia II - 26-04-2017 - RDocumento11 pagine30 - Anatomia II - 26-04-2017 - RSandroNessuna valutazione finora
- 10 - Traumatologia Della Colonna - 25-03-14Documento4 pagine10 - Traumatologia Della Colonna - 25-03-14Ilaria PuttiniNessuna valutazione finora
- La Caviglia Ed Il Piede Nello SportDocumento5 pagineLa Caviglia Ed Il Piede Nello SportCipriano D'AmbrosioNessuna valutazione finora
- Lezione II-10 Endocrino 04-04-11Documento91 pagineLezione II-10 Endocrino 04-04-11Roberta RossiNessuna valutazione finora
- Patologie Specifiche Del TennisDocumento1 paginaPatologie Specifiche Del TennisTurrix 1sNessuna valutazione finora
- Messaggeri ChimiciDocumento30 pagineMessaggeri ChimiciMarynaNessuna valutazione finora
- Lesioni MuscolariDocumento12 pagineLesioni MuscolariFrancoNessuna valutazione finora
- Iperuricemia e Gotta CorrettaDocumento7 pagineIperuricemia e Gotta CorrettaDavideNessuna valutazione finora
- Paratiroidi 2006Documento49 pagineParatiroidi 2006api-3733154100% (1)
- 11 Anatomia 20-03-14Documento10 pagine11 Anatomia 20-03-14SandroNessuna valutazione finora
- I Sette Livelli Reichiani e La Psicologia OlisticaDocumento3 pagineI Sette Livelli Reichiani e La Psicologia OlisticaMigliorino CarmeloNessuna valutazione finora
- 22 - Anatomia II - 22.03.2017 - RDocumento8 pagine22 - Anatomia II - 22.03.2017 - RSandroNessuna valutazione finora
- Paramorfismi e DismorfismiDocumento20 pagineParamorfismi e DismorfismiTiziana FioreNessuna valutazione finora
- Sinapsi e NeurotrasmettitoriDocumento45 pagineSinapsi e NeurotrasmettitoriNicola Laurora0% (1)
- 4 Lezione - La Cute e I Suoi AnnessiDocumento24 pagine4 Lezione - La Cute e I Suoi AnnessiAlbert stefan PaulencoNessuna valutazione finora
- Patologie Dell'Esofago - Patologia Chirurgica (Università Di Siena)Documento37 paginePatologie Dell'Esofago - Patologia Chirurgica (Università Di Siena)CARLOTTA IADANZANessuna valutazione finora
- Anatomia 12-03-18Documento16 pagineAnatomia 12-03-18arancinetopNessuna valutazione finora
- Apparato DigerenteDocumento5 pagineApparato DigerenteMaria TeresaNessuna valutazione finora
- 26 - Anatomia II - 30-03-2017 (R)Documento10 pagine26 - Anatomia II - 30-03-2017 (R)SandroNessuna valutazione finora
- 43-Anatomia II-14.04.2016 - Fegato, Pancreas e MilzaDocumento7 pagine43-Anatomia II-14.04.2016 - Fegato, Pancreas e MilzaSandroNessuna valutazione finora
- Ricerche Sulle Nuove Applicazioni Delle Piante Officinali in FitocosmesiDocumento4 pagineRicerche Sulle Nuove Applicazioni Delle Piante Officinali in FitocosmesiRenato BruniNessuna valutazione finora
- Triangolo Femorale o Di ScarpaDocumento2 pagineTriangolo Femorale o Di ScarpaVincenzo GiulianoNessuna valutazione finora
- Equiseto ArvenseDocumento61 pagineEquiseto ArvensesardegnamovieNessuna valutazione finora
- Viscerale Lezione 1 Parte 2Documento13 pagineViscerale Lezione 1 Parte 2Antonio GarNessuna valutazione finora
- Diaframma PDFDocumento2 pagineDiaframma PDFPaoly PalmaNessuna valutazione finora
- 13 - Immunologia-26.04.2017Documento11 pagine13 - Immunologia-26.04.2017SandroNessuna valutazione finora
- 31-Anatomia II-03.03.2016-Topografia Anatomica Del CuoreDocumento6 pagine31-Anatomia II-03.03.2016-Topografia Anatomica Del CuoreSandroNessuna valutazione finora
- SV - 09 - Malattie Del Metabolismo - Gotta, Obesità e Diabete in Gravidanza - Lapolla - 31-03-2022Documento15 pagineSV - 09 - Malattie Del Metabolismo - Gotta, Obesità e Diabete in Gravidanza - Lapolla - 31-03-2022Michele GiordanoNessuna valutazione finora
- Appunti Di Anatomia Umana v. 2020Documento17 pagineAppunti Di Anatomia Umana v. 2020CiroNessuna valutazione finora
- Gli Shen Organi e VisceriDocumento14 pagineGli Shen Organi e VisceriEr AimNessuna valutazione finora
- PancreasDocumento34 paginePancreasy666t5bvbfgNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- Muscolo Schel - Parte1Documento34 pagineMuscolo Schel - Parte1Francesco MalavoltaNessuna valutazione finora
- Case-Sindrome-Scaleni Sindrome Stretto Toracico Prima CostaDocumento5 pagineCase-Sindrome-Scaleni Sindrome Stretto Toracico Prima CostaAlessandroVolpeNessuna valutazione finora
- 20 - Anatomia II - 16.03.2017 - RDocumento10 pagine20 - Anatomia II - 16.03.2017 - RSandroNessuna valutazione finora
- La Teoria Dei Cinque Elementi PDFDocumento20 pagineLa Teoria Dei Cinque Elementi PDFr4dn4vNessuna valutazione finora
- Istologia - ConnettivoDocumento46 pagineIstologia - ConnettivoRosario GrassoNessuna valutazione finora
- 5-Stenosi MitralicaDocumento8 pagine5-Stenosi MitralicaScribdNixNessuna valutazione finora
- Allenamento Muscoli Posteriori Coscia PDFDocumento7 pagineAllenamento Muscoli Posteriori Coscia PDFElisa BettaNessuna valutazione finora
- Anatomia e Fisiologia Del Sistema MuscolareDocumento36 pagineAnatomia e Fisiologia Del Sistema MuscolareFrancescaNessuna valutazione finora
- Malattie Del Sistema Endocrino e MetabolismoDocumento55 pagineMalattie Del Sistema Endocrino e MetabolismoOr MillerNessuna valutazione finora
- Curare I Dolori Con La Riflessologia PlantareDocumento16 pagineCurare I Dolori Con La Riflessologia PlantareclekupreNessuna valutazione finora
- UntitledDocumento23 pagineUntitledMartina ZuccariniNessuna valutazione finora
- Sillabario Di Radioprotezione: Prof. Giovanni Luca GravinaDocumento24 pagineSillabario Di Radioprotezione: Prof. Giovanni Luca GravinaMartina ZuccariniNessuna valutazione finora
- Radioprotezione in Radioterapia: Prof. Giovanni Luca GravinaDocumento43 pagineRadioprotezione in Radioterapia: Prof. Giovanni Luca GravinaMartina ZuccariniNessuna valutazione finora
- UntitledDocumento29 pagineUntitledMartina ZuccariniNessuna valutazione finora
- Lezione 14Documento12 pagineLezione 14simoNessuna valutazione finora
- Patologia GeneraleDocumento179 paginePatologia GeneraleChiara LepriNessuna valutazione finora
- Parete Cellulare BattericaDocumento3 pagineParete Cellulare BattericaJoel NiciakNessuna valutazione finora
- 14-Emostasi PrimariaDocumento43 pagine14-Emostasi PrimariaSpinelli PietroNessuna valutazione finora
- LEZIONE 8 Ciclo Di Krebs e Fosforilazione OssidativaDocumento53 pagineLEZIONE 8 Ciclo Di Krebs e Fosforilazione OssidativaMaegazine EnizageamNessuna valutazione finora
- La Parete Cellulare-Prima ParteDocumento40 pagineLa Parete Cellulare-Prima PartenomeeeeNessuna valutazione finora
- Citologia-A. Di Golgi LisosomiDocumento30 pagineCitologia-A. Di Golgi LisosomiHamidreza DeldadehNessuna valutazione finora
- Bio Molecolare TotaleDocumento13 pagineBio Molecolare TotaleCarlo SavioliNessuna valutazione finora
- Medicina Olistica Energetica: Guarire percorrendo una strada alternativa alla scienzaDa EverandMedicina Olistica Energetica: Guarire percorrendo una strada alternativa alla scienzaValutazione: 5 su 5 stelle5/5 (1)
- Nuovo Esame per Esperto in Gestione dell'Energia - Settore Civile: Test e temi di esame svolti per sostenere l’esame di Esperto in Gestione dell’Energia del Settore CivileDa EverandNuovo Esame per Esperto in Gestione dell'Energia - Settore Civile: Test e temi di esame svolti per sostenere l’esame di Esperto in Gestione dell’Energia del Settore CivileNessuna valutazione finora
- Anatomia della Coscienza Quantica: La fisica dell'auto-guarigioneDa EverandAnatomia della Coscienza Quantica: La fisica dell'auto-guarigioneValutazione: 4.5 su 5 stelle4.5/5 (3)
- La medicina di santa Ildegarda: Per tutti i giorni e per tutta la famiglia.Da EverandLa medicina di santa Ildegarda: Per tutti i giorni e per tutta la famiglia.Valutazione: 5 su 5 stelle5/5 (2)
- Chimica Occulta: Indagine nella struttura occulta degli atomiDa EverandChimica Occulta: Indagine nella struttura occulta degli atomiNessuna valutazione finora
- Elettrotecnica Generale: Circuiti Elettrici in Regime StazionarioDa EverandElettrotecnica Generale: Circuiti Elettrici in Regime StazionarioNessuna valutazione finora
- La comunicazione strategica nelle professioni sanitarieDa EverandLa comunicazione strategica nelle professioni sanitarieNessuna valutazione finora
- Biologia quantistica e Cronobiologia fasciale osteopaticaDa EverandBiologia quantistica e Cronobiologia fasciale osteopaticaNessuna valutazione finora
- Approccio ai disordini dell'EAB: L'Equilibrio Acido-Base nella pratica clinicaDa EverandApproccio ai disordini dell'EAB: L'Equilibrio Acido-Base nella pratica clinicaNessuna valutazione finora
- REIKI TRADIZIONALE GIAPPONESE primo e secondo livelloDa EverandREIKI TRADIZIONALE GIAPPONESE primo e secondo livelloNessuna valutazione finora
- Biologia quantica: Viaggio ai confini della guarigioneDa EverandBiologia quantica: Viaggio ai confini della guarigioneNessuna valutazione finora
- Elettrotecnica | Passo dopo passo: Fondamenti, componenti & circuiti spiegati per principiantiDa EverandElettrotecnica | Passo dopo passo: Fondamenti, componenti & circuiti spiegati per principiantiNessuna valutazione finora
- Morfologia Dentale: La forma come fondamenta della modellazione sequenziale dall'analogico al digitaleDa EverandMorfologia Dentale: La forma come fondamenta della modellazione sequenziale dall'analogico al digitaleNessuna valutazione finora
- Anatomia della Guarigione: I sette principi della Nuova Medicina IntegrataDa EverandAnatomia della Guarigione: I sette principi della Nuova Medicina IntegrataValutazione: 5 su 5 stelle5/5 (2)
- Simbologia Massonica Vol.I: Simbologia e Tradizione esoterica liberomuratoriaDa EverandSimbologia Massonica Vol.I: Simbologia e Tradizione esoterica liberomuratoriaNessuna valutazione finora
- Il Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoDa EverandIl Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoNessuna valutazione finora
- Razza e Storia e altri studi di AntropologiaDa EverandRazza e Storia e altri studi di AntropologiaValutazione: 5 su 5 stelle5/5 (2)