Potrebbero piacerti anche
- Membrana PlasmaticaDocumento11 pagineMembrana PlasmaticaRossella MeolaNessuna valutazione finora
- Morte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cDa EverandMorte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cNessuna valutazione finora
- Struttura e Funzioni Della Membrana PlasmaticaDocumento4 pagineStruttura e Funzioni Della Membrana Plasmaticagiulia petraccaNessuna valutazione finora
- Biologia 3 - Membrana PlasmaticaDocumento10 pagineBiologia 3 - Membrana PlasmaticaChiara MontanoNessuna valutazione finora
- La Membrana Cellulare e Le Membrane InterneDocumento5 pagineLa Membrana Cellulare e Le Membrane InterneIrene MazzolaNessuna valutazione finora
- 2.membrane CellulariDocumento25 pagine2.membrane CellulariCalogero TodaroNessuna valutazione finora
- Lezione 2 - Biologia Cellulare - 23 - 10Documento17 pagineLezione 2 - Biologia Cellulare - 23 - 10beatris.zaharia04Nessuna valutazione finora
- La Membrana CellulareDocumento4 pagineLa Membrana CellulareMiriamPellitteriNessuna valutazione finora
- 4-Modello A Mosaico Fluido e Fluidità MembraneDocumento2 pagine4-Modello A Mosaico Fluido e Fluidità MembranewagamifNessuna valutazione finora
- Fisiologia SbobbineDocumento2 pagineFisiologia Sbobbinechantal.dilizioNessuna valutazione finora
- Sistesis de LipidosDocumento2 pagineSistesis de LipidosbertasierramaquedaNessuna valutazione finora
- 4 Le Membrane CellulariDocumento19 pagine4 Le Membrane CellulariAlessia MaraNessuna valutazione finora
- Fisiologia - )Documento270 pagineFisiologia - )Ilaria ZeddaNessuna valutazione finora
- Appunti Citologia e IstologiaDocumento73 pagineAppunti Citologia e IstologiaAntonio LambresaNessuna valutazione finora
- Macromolecole - Citologia e IstologiaDocumento42 pagineMacromolecole - Citologia e IstologiasubzeroNessuna valutazione finora
- BIOAPP - TEC - 18 Lez 3 Le Membrane CellulariDocumento36 pagineBIOAPP - TEC - 18 Lez 3 Le Membrane CellulariDomenico ZucaroNessuna valutazione finora
- Il Modello A Mosaico Fluido-ProteineDocumento1 paginaIl Modello A Mosaico Fluido-ProteineMariella SiervoNessuna valutazione finora
- CitologiaDocumento6 pagineCitologiajobronyNessuna valutazione finora
- La CellulaDocumento10 pagineLa CellulaAngelica CoppolaNessuna valutazione finora
- Relazione Sulla Cellula Eucariote AnimaleDocumento8 pagineRelazione Sulla Cellula Eucariote AnimaleMarco Pio SessaNessuna valutazione finora
- Appunti Di Fisiologia UltimiDocumento51 pagineAppunti Di Fisiologia UltimiLuisa Ghoul Di SciascioNessuna valutazione finora
- Membrane BiologicheDocumento81 pagineMembrane BiologicheannaNessuna valutazione finora
- CitologiaDocumento53 pagineCitologianameNessuna valutazione finora
- Citologia IstologiaDocumento85 pagineCitologia IstologiaFrancesco GualdiNessuna valutazione finora
- Ist Ed Embr Lezione 3Documento34 pagineIst Ed Embr Lezione 3sereve02Nessuna valutazione finora
- I LisosomiDocumento1 paginaI LisosomiCosso GiuliaNessuna valutazione finora
- 3 Trasporti Di MembranaDocumento14 pagine3 Trasporti Di MembranalorellaNessuna valutazione finora
- Plastidi FATTODocumento44 paginePlastidi FATTOArianna LandFNessuna valutazione finora
- Cap. 7 Membrana PlasmaticaDocumento6 pagineCap. 7 Membrana PlasmaticaRosemarie ScaliNessuna valutazione finora
- Riassunto FisiologiaDocumento31 pagineRiassunto FisiologiaHibino TsubakiNessuna valutazione finora
- MembraneDocumento5 pagineMembraneAlessia ShadowhuntersNessuna valutazione finora
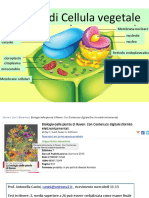
- Cellula VegetaleDocumento58 pagineCellula VegetaleEleonora AngelocolaNessuna valutazione finora
- Appunti BiologiaDocumento12 pagineAppunti BiologiaAlessia PolitoNessuna valutazione finora
- Membrana Plasmatica 1Documento3 pagineMembrana Plasmatica 1Lucas RomeroNessuna valutazione finora
- La Cellula Al LavoroDocumento3 pagineLa Cellula Al LavoroGiovanni MichielinNessuna valutazione finora
- Materia ViventeDocumento164 pagineMateria ViventeRosi De LucaNessuna valutazione finora
- Biologia CellulareDocumento6 pagineBiologia CellulareClaudia BertenniNessuna valutazione finora
- Materia ViventeDocumento5 pagineMateria ViventeRosi De LucaNessuna valutazione finora
- Citologia-Generalità e MembranaDocumento32 pagineCitologia-Generalità e MembranaHamidreza DeldadehNessuna valutazione finora
- Fisiologia 1 - TrasportiDocumento19 pagineFisiologia 1 - TrasportiAnna De Simone100% (1)
- Lezione 15Documento5 pagineLezione 15bertasierramaquedaNessuna valutazione finora
- 1163 - La Membrana Plasmatica PDFDocumento8 pagine1163 - La Membrana Plasmatica PDFDiego Flores EgüezNessuna valutazione finora
- Trasporto PassivoDocumento3 pagineTrasporto Passivogiulia petraccaNessuna valutazione finora
- Le BiomolecoleDocumento14 pagineLe BiomolecoleAlessandra TessitoreNessuna valutazione finora
- Capitolo 2 I Batteri MicrobiologiaDocumento49 pagineCapitolo 2 I Batteri MicrobiologiaClaudia Di finizioNessuna valutazione finora
- La Cellula - WikibookDocumento107 pagineLa Cellula - WikibookMichele Scipioni100% (1)
- Cap 1Documento2 pagineCap 1ivana elefanteNessuna valutazione finora
- RIASSUNTO Programma Peresame - BiologiaDocumento23 pagineRIASSUNTO Programma Peresame - BiologiaSilvia GrossoNessuna valutazione finora
- Lez 12 CHIM ORG E BIOCH 2023Documento31 pagineLez 12 CHIM ORG E BIOCH 2023mallnitz04Nessuna valutazione finora
- Lezione 3 - Biologia Cellulare - 24 - 10Documento14 pagineLezione 3 - Biologia Cellulare - 24 - 10beatris.zaharia04Nessuna valutazione finora
- 07l03 Membrana CellDocumento3 pagine07l03 Membrana Cellracie (boveerica)Nessuna valutazione finora
- LA CellulaDocumento3 pagineLA CellulaCosso GiuliaNessuna valutazione finora
- BIOCHIMICA - SiliprandiDocumento50 pagineBIOCHIMICA - SiliprandiinfernuxNessuna valutazione finora
- Citologia IstologiaDocumento46 pagineCitologia IstologiaElisa TribuianiNessuna valutazione finora
- Botanica Cap 6Documento2 pagineBotanica Cap 6ivana elefanteNessuna valutazione finora
- Domande MadedduDocumento13 pagineDomande MadedduClaire ColomboNessuna valutazione finora
- Fisiologia UmanaDocumento68 pagineFisiologia UmanaMariarita RiccardiNessuna valutazione finora
- Ricerca Struttura e Funzioni Della Membrana PlasmaticaDocumento11 pagineRicerca Struttura e Funzioni Della Membrana Plasmaticafrancesco.fanesi01Nessuna valutazione finora
- Lezione 12 Colesterolo PDFDocumento61 pagineLezione 12 Colesterolo PDFpaolo68audisioNessuna valutazione finora
- Appunti Di BotanicaDocumento169 pagineAppunti Di BotanicaAlfonso GrimaldiNessuna valutazione finora
- BiochimicaDocumento28 pagineBiochimicahayhaNessuna valutazione finora
- Lezione 14Documento12 pagineLezione 14simoNessuna valutazione finora
- Biochimica Rec 14Documento5 pagineBiochimica Rec 14hayhoNessuna valutazione finora
- BiochimicaDocumento28 pagineBiochimicahayhaNessuna valutazione finora
- Saponificazione EtAcDocumento5 pagineSaponificazione EtAcFabioNessuna valutazione finora
- Domande Di Teoria Orale Fisica TecnicaDocumento5 pagineDomande Di Teoria Orale Fisica TecnicaAnna ScaburriNessuna valutazione finora
- La Conducibilità ElettricaDocumento2 pagineLa Conducibilità ElettricaPalermo Scienza100% (2)
- Corso Di Tecnologia Meccanica - Mod 5.2 Lavorazioni Non ConvenzionaliDocumento47 pagineCorso Di Tecnologia Meccanica - Mod 5.2 Lavorazioni Non ConvenzionaliDario CaparròzNessuna valutazione finora
- Chimica: Cos'è L'atomo?Documento37 pagineChimica: Cos'è L'atomo?Van MorrisonNessuna valutazione finora
- Le Tattiche del Cambiamento - Manuale di Psicoterapia StrategicaDa EverandLe Tattiche del Cambiamento - Manuale di Psicoterapia StrategicaValutazione: 5 su 5 stelle5/5 (1)
- Morfologia Dentale: La forma come fondamenta della modellazione sequenziale dall'analogico al digitaleDa EverandMorfologia Dentale: La forma come fondamenta della modellazione sequenziale dall'analogico al digitaleNessuna valutazione finora
- Agnihotri - I Custodi del Fuoco: Il Sentiero di Guarigione del Guerriero del FuocoDa EverandAgnihotri - I Custodi del Fuoco: Il Sentiero di Guarigione del Guerriero del FuocoNessuna valutazione finora
- Chimica Occulta: Indagine nella struttura occulta degli atomiDa EverandChimica Occulta: Indagine nella struttura occulta degli atomiNessuna valutazione finora
- Nuovo Esame per Esperto in Gestione dell'Energia - Settore Civile: Test e temi di esame svolti per sostenere l’esame di Esperto in Gestione dell’Energia del Settore CivileDa EverandNuovo Esame per Esperto in Gestione dell'Energia - Settore Civile: Test e temi di esame svolti per sostenere l’esame di Esperto in Gestione dell’Energia del Settore CivileNessuna valutazione finora
- Elettrotecnica | Passo dopo passo: Fondamenti, componenti & circuiti spiegati per principiantiDa EverandElettrotecnica | Passo dopo passo: Fondamenti, componenti & circuiti spiegati per principiantiNessuna valutazione finora
- Il Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoDa EverandIl Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoNessuna valutazione finora
- Umberto Galimberti Eugenio Borgna Un luminare miopeDa EverandUmberto Galimberti Eugenio Borgna Un luminare miopeNessuna valutazione finora
- Elaborato finale dell’attività di tirocinio. Progettazione di un percorso educativo-didattico in: sicurezza stradale a carattere inclusivoDa EverandElaborato finale dell’attività di tirocinio. Progettazione di un percorso educativo-didattico in: sicurezza stradale a carattere inclusivoNessuna valutazione finora
- REIKI TRADIZIONALE GIAPPONESE primo e secondo livelloDa EverandREIKI TRADIZIONALE GIAPPONESE primo e secondo livelloNessuna valutazione finora
- Manipolazione: Padroneggia tecniche di persuasione altamente efficaci, controllo mentale e influenza emotiva utilizzando la psicologia oscura, come analizzare le persone, il linguaggio del corpo, i segreti della PNL e dell'ipnosi!Da EverandManipolazione: Padroneggia tecniche di persuasione altamente efficaci, controllo mentale e influenza emotiva utilizzando la psicologia oscura, come analizzare le persone, il linguaggio del corpo, i segreti della PNL e dell'ipnosi!Valutazione: 5 su 5 stelle5/5 (4)
- Depressione, ansia e panico: la terapia psicologicaDa EverandDepressione, ansia e panico: la terapia psicologicaNessuna valutazione finora
- Esercizi svolti di Matematica: Geometria AnaliticaDa EverandEsercizi svolti di Matematica: Geometria AnaliticaNessuna valutazione finora
- Come addormentarsi: Le guide rapide ed efficaci per chi ha poco tempoDa EverandCome addormentarsi: Le guide rapide ed efficaci per chi ha poco tempoNessuna valutazione finora
- Medicina Olistica Energetica: Guarire percorrendo una strada alternativa alla scienzaDa EverandMedicina Olistica Energetica: Guarire percorrendo una strada alternativa alla scienzaValutazione: 5 su 5 stelle5/5 (1)
- Come riconoscere un narcisista: Conoscere il narcisismoDa EverandCome riconoscere un narcisista: Conoscere il narcisismoNessuna valutazione finora
- Il Libricino della Felicità: come liberarsi dalle zavorre e raggiungere i propri obiettiviDa EverandIl Libricino della Felicità: come liberarsi dalle zavorre e raggiungere i propri obiettiviValutazione: 5 su 5 stelle5/5 (3)