Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
Appunti Di Neurofisiologia
Caricato da
SaraTitolo originale
Copyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
Appunti Di Neurofisiologia
Caricato da
SaraCopyright:
Formati disponibili
appunti di neurofisiologia
Neuroscienze
Università degli Studi di Sassari (UNISS)
90 pag.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
INTRODUZIONE AL SNC
L’encefalo è costituito dal cervello (diviso in telencefalo e diencefalo), dal tronco encefalico (bulbo,
ponte e mesencefalo) e dal cervelletto.
Il primo a visualizzare i neuroni fu Golgi. creò un nuovo metodo di colorazione, attraverso il quale
però non si colorano tutti i neuroni. Inoltre Golgi era convinto che tutti i neuroni fossero collegati
fisicamente tra di loro (teoria reticolare o sinciziale), quindi che ci fosse un’enorme rete di cellule
tutte attaccate le une alle altre. Cajal sviluppò la neuron doctrine; ossia nega la teoria reticolare e
afferma che i neuroni sono unità singole che comunicano con gli altri neuroni attraverso punti di
contatto e che il neurone sia l’unità funzionale del SNC. Grazie a Golgi sappiamo comunque che
esistono tanti tipi differenti di neuroni, ma derivano tutti dalla stessa cellula attraverso la
neurogenesi; la forma ci permette di distinguerli in diverse famiglie; alcune famiglie neuronali
sono più presenti di altre nel nostro encefalo (neurone piramidale corteccia cerebrale, cellule di
Golgi corteccia cerebellare).
NEURONI: I neuroni sono semipermeabili, solo alcuni ioni possono attraversare i canali presenti
sulla membrana. Ha delle precise caratteristiche:
-Genera il Potenziale d’Azione
-Risponde a degli stimoli
-Integra le informazioni ricevute dagli altri neuroni -Trasporta un segnale bioelettrico
-Comunica con altre cellule
-Si adatta plasticamente
In un neurone individuiamo quattro aree principali:
-soma: parte centrale e zona metabolica del
neurone, si tratta del corpo cellulare;
-dendriti: processo neuronale spesso e arborizzato
che si trova vicino al soma; i dendriti di molti neuroni
sono caratterizzati da piccole protrusioni, le spine
dendritiche, anch’esse coinvolte nel trasferimento
dell’informazione tra cellule;
-assone: processo neuronale lungo e sottile che si
estende molto oltre il soma e in alcuni casi possiede una guaina mielinica (il suo danneggiamento
comporta sclerosi multipla); all’estremità degli assoni ci sono i terminali presinaptici, strutture
specializzate che partecipano al trasferimento di informazioni tra neuroni.
-Le terminazioni assoniche: area in cui il segnale (trasportato dall’assone) viene rilasciato
(attraverso la sinapsi) per confluire verso il neurone successivo.
SINAPSI:
La sinapsi rappresenta la metodologia comunicativa del neurone. Esistono due tipi di sinapsi,
quella chimica e quella elettrica. Entrambe, in base alla loro localizzazione geografica, possono
essere definite assodendritiche, assosomatiche o assosoniche.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
- Le sinapsi chimiche (maggiormente presenti all’interno del S.N.C) hanno la peculiarita’ di
trasmettere una o piu’ vescicole di neurotrasmettitore (sostanza chimica eccitatoria od inibitrice)
lungo l’assone. La vescicola viene rilasciata (dopo aver raggiunto l’ area terminale dell’assone
della cellula pre-sinaptica) nello spazio sinaptico (ossia nello spazio bianco dell’immagine).
Sara’ compito della cellula post-sinaptica recepire il neurotrasmettitore attraverso dei recettori di
riferimento (di cui parleremo in seguito).
Caratteristiche S. chimica:
-Amplifica il segnale
-Elabora il segnale per sommazione (una maggiore
rilascio di vescicole contenenti
neurotrasmettitori daranno vita un messaggio piu’
“forte” e viceversa)
-Bassa velocita’ di trasmissione (se paragonata a
quella elettrica)
-Affaticabilita’ (se le vescicole terminano serve del
tempo perche’ si riformulano) -Vulnerabilita’ (una
sostanza diversa dal neurotrasmettitore puo’
“assumere le sembianze”
del neurotrasmettitore ed essere recepito dal recettore di riferimento al suo posto).
- Le sinapsi elettriche (maggiormente presenti, ad esempio, nel muscolo cardiaco) producono
un impulso di tipo elettrico (piu’ semplicemente un flusso
ionico) che si sprigiona grazie a dei canali - definiti gap junctions - i quali pongono in
comunicazione (ed uniscono) la cellula post e quella pre-sinaptica.
Caratteristiche S. Elettriche:
-Semplicita’ strutturale
-Elevata velocita’ di trasmissione
-Basso consumo energetico
-Bidirezionalita’ del segnale (al contrario di quelle chimiche) -Staticita’ (non cambiano le
loro “performance”)
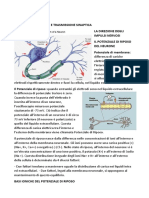
Il Potenziale d’Azione:
Il neurone ha un proprio Potenziale di Membrana: una differenza di potenziale elettrico tra
l’interno e l’esterno della cellula. Il potenziale d’azione è un segnale bioelettrico, l’unità
elementare degli impulsi nervosi che gli assoni usano per trasferire informazioni anche a lunga
distanza, mostrano la proprietà del tutto o nulla. I neuroni usano la frequenza dei potenziali
d’azione per trasferire l’intensità dei segnali che devono trasmettere.
Il neurone, a riposo, ha un potenziale (precedentemente definito potenziale di riposo) di circa
-70mV. Quando si manifesta un potenziale d’azione il neurone inverte la polarizzazione della
propria membrana che passa da circa -70mV a circa +40mV (definiremo questa fase
“depolarizzazione”) per poi – durante una seconda fase - scendere addirittura al di sotto del
valore di riposo, fino a circa -80mV (definiremo questa fase “iperpolarizzazione” o
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
“ripolarizzazione”) per poi – infine – tornare lentamente al proprio potenziale di riposo originario
(-70mV).
e’ essenziale comprendere il P.A anche da un punto di vista biochimico. E’ quindi inevitabile
proporre alcuni concetti di base senza i quali risulterebbe impossibile capire gli argomenti
successivi:
Gli ioni coinvolti nel P.A sono gli NA+ e gli K+
I canali ionici sono delle proteine che - penetrate all’interno della membrana cellulare-
fungono da “varco” e consentono il passaggio degli ioni tra l’ area intracellulare e quella
extracellulare (e viceversa).
Ogni ione ha il proprio canale ionico di riferimento (gli NA+ non possono circolare nei canali
ionici K+ e viceversa)
I canali ionici si aprono e si chiudono in risposta a dei segnali:
-Segnali elettrici (canali voltaggio dipendenti, Regolati da variazioni di voltaggio);
-Segnali chimici (canali ligando dipendenti, regolati da neurotrasmettitori chimici);
-Segnali meccanici (canali meccanico dipendenti, regolati da pressione o stiramento)
Le pompe ioniche consentono il passaggio di alcuni ioni ma NON sono dei canali ionici,
hanno infatti delle caratteristiche differenti: consumano ATP e gestiscono un flusso ionico da
100 a 100.000 volte inferiore a quello dei canali ionici.
Esistono dei canali ionici PASSIVI (che sostanzialmente sono sempre aperti) importantissimi
soprattutto per il mantenimento del gradiente di concentrazione.
- Da un punto di vista biochimico il P.A agisce in questo modo:
La cellula non riceve afferenze (e’ a riposo), il suo potenziale e’ di -70mV, i canali passivi del
NA+ sono aperti e la pompa sodio/potassio trasporta NA+ all’esterno della cellula e K+
all’interno della cellula.
I canali voltaggio dipendenti sono chiusi. Quelli passivi del K+ aperti.
Delle afferenze (stimoli) possono portare ad una variazione del numero degli ioni all’interno
della cellula. Ciò causa una depolarizzazione della membrana.
Se le afferenze portano al raggiungimento del potenziale soglia di circa -50mV, si aprono i
canali voltaggio dipendenti del sodio
E sodio entra all'interno della cellula portandola a circa +40 mV. L’inversione della Carica
consente Inoltre l'apertura dei canali voltaggio dipendente K+ (I quali sono però più lenti ad
aprirsi di quelli del NA+).
I canali NA+ si inattivano.
La membrana si iperpolarizza con l’ uscita del K
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
A causa della lentezza dell’apertura e della chiusura dei canali del K+ esso continua ad
uscire fino a circa -80mV e a pompa NA+ K+ (pompa sodio/potassio) ristabilisce la
condizione ionica iniziale.
Dal punto 3 fino a circa meta’ del punto 6 la cellula e’ in una condizione di refrattarieta’
assoluta. Cio’ significa che non puo’ rispondere ad alcuno stimolo (proprio a causa del fatto
che i canali del NA+ sono inattivi).
Dalla meta’ del punto 6 alla fine del punto 8 e’ invece in refrattarieta’ relativa. Cio’ significa che
il neurone e’ diventato nuovamente eccitabile ma per raggiungere il valore di soglia (che
convenzionalmente abbiamo indicato in
-50mV) e per avere la possibilita’ di sparare un altro P.A , dovra’ subire uno stimolo maggiore.
-Caratteristiche del Potenziale d’Azione:
- E’ evocato da una depolarizzazione della membrana
- Ha un valore di potenziale soglia oltre il quale il P.A si manifesta - E’ un fenomeno attivo e
che si autorigenera
- E’ caratterizzato da dei periodi di refrattarieta’
- E’ una risposta del tipo “ tutto o nulla” (o spara o non spara)
- E’ una risposta stereotipata
- Trasporta informazioni
POTENZIALE GRADUATO
Il potenziale graduato o potenziale locale è la continua variazione del potenziale di membrana
rispetto al potenziale d’azione (esempio di potenziale graduato è il potenziale sinaptico, prodotto a
livello dei siti postsinaptici in risposta al rilascio del neurotrasmettitore da parte del terminale
presinaptico.
I potenziali graduati invece:
-Si muovono senza una direzione obbligatoria
-Sono influenzati dalla struttura anatomica della membrana
-La distanza dal soma ne determina l’influenza che ha sulla cellula stessa
-Possono essere sia “positivi” che “negativi”
-Possono avere un’ onda (graficamente parlando) piu’ lunga e bassa o piu’ alta e corta, a
seconda della propria potenza
- Si manifestano tendenzialmente nel’ area dendritica
-Possono soprattutto SOMMARSI TRA LORO TEMPORALMENTE (in base cioe’ alla
propria cadenza temporale) ED A LIVELLO SPAZIALE (in base cioe’ alla distanza dei
singoli potenziali sprigionati).
Tra l’ambiente interno ed esterno della cellula abbiamo la membrana plasmatica, la cui parte
idrofila è rivolta verso l’esterno mentre le code idrofobe (di acidi grassi) sono rivolte all’interno (es.
barriera ematoencefalica). La membrana delle cellule umane è quindi formata da un doppio strato
fosfolipidico impermeabile agli ioni, quindi le molecole dotate di carica elettrica non sono capaci di
attraversarla, motivo per cui esistono le proteine di membrana, che formano dei canali che mettono
in comunicazione l’interno e l’esterno della cellula.
Esistono diversi tipi di canali, ma la caratteristica comune è che sono tutti ionici, sono tutti proteine
transmembrana che attraversano il doppio strato fosfolipidico, sostenendo un poro che permette il
passaggio di alcuni ioni.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
I due mondi separati dalla membrana plasmatica sono differenti, ma non per la loro composizione:
gli ioni sono gli stessi, cambia la loro concentrazione.
Ambiente intracellulare: Na+ 50mM; K+ 400mM; Cl- 40mM; Ca2+ 10−4mM Ambiente extracellulare:
Na+ 440mM; K+ 20mM; Cl- 560mM; Ca2+ 10mM
Il potenziale di membrana non è dovuto da questa differenza quantitativa, bensì dal fatto che gli
ioni si possono muovere.
Le cellule gliali si riproducono anche nell’età adulta, a differenza dei neuroni hanno funzione di
sostegno meccanico e isolamento elettrico (oligodendrociti e cellule di Schwann ) hanno
funzione di “spazzini”: fagocitosi di microbi e detriti cellulari (microglia) produzione del liquido
cefalorachidiano (cellule ependimali) guidano la rigenerazione delle fibre danneggiate
CELLULE ECCITABILI
In condizioni di riposo la membrana delle cellule eccitabili è polarizzata (potenziale di riposo).
Quando la membrana viene eccitata si assiste ad una modifica del suo stato elettrico
(potenziale
d’azione) che ritorna poi alle condizioni iniziali (potenziale di riposo). La polarizzazione della
membrana trae origine da una diversa distribuzione delle cariche (ioni) a cavallo della
membrana e da un differente flusso delle stesse attraverso la membrana medesima. Il flusso
degli ioni attraverso la membrana dipende da 4 fattori:
1. Diversa concentrazione degli ioni ai due lati della membrana;
2. Diversa permeabilità della
membrana agli ioni ;
3. Separazione transmembranari delle cariche;
4. Trasporto attivo di alcuni ioni (es. Na+ e K+ ) attraverso la membrana
NEUROTRASMETTITORE
Un neurotrasmettitore è una sostanza chimica in grado di legarsi a recettori specifici presenti
sulla membrana della cellula bersaglio.
Liberato generalmente dalle terminazioni presinaptiche, rigonfiamenti specializzati dell’assone
che, in genere, contengono da 100 a 200 vescicole sinaptiche, ciascuna delle quali contiene
parecchie migliaia di molecole di neurotrasmettitore.
Le vescicole si accumulano in zone della membrana specializzate per il loro rilascio, dette zone
attive.
AMINOACIDI AMMINE
Glutammato: principale Nt eccitatorio centrale Noradrenalina: sistema nervoso autonomo:
simpatico post gangliare,sist nerv centr
GABA: principale Nt inibitorio centrale Dopamina, es. gangli della base
Glicina: principale Nt inibitorio spinale Serotonina(5-HT) es. Nuclei del rafe,
ipotalamo
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Sistema di rilascio del neurotrasmettitore
Nella sinapsi elettrica il segnale di tipo bioelettrico passa da una cellula all’altra, restando segnale
bioelettrico. Nella sinapsi chimica, invece, il segnale bioelettrico viene trasformato quando arriva
alla parte presinaptica in un segnale chimico, il neurotrasmettitore appunto, che viene rilasciato e
che arriva nella cellula postsinaptica ridiventando segnale bioelettrico, diverso da quello originale:
se prima si trattava di potenziale d’azione, adesso è un potenziale graduato. Le tappe della
neurotrasmissione sono:
⤿arrivo del potenziale d’azione al bottone presinaptico
⤿apertura dei canali Ca2+ voltaggio dipendenti (cambiano il loro stato a seconda del voltaggio di
membrana)
⤿il Ca2+ fa fondere le vescicole sinaptiche con la membrana plasmatica e il neurotrasmettitore
viene liberato in uno spazio tra due cellule, lo spazio sinaptico (più calcio entra, più vescicole
vengono liberate) interagisce un recettore della cellula postsinaptica il neurotrasmettitore si lega al
recettore sinaptotagmina e quest’ultimo cambia il suo stato (ligando dipendente) ciò apre un flusso
ionico: viene indotto un potenziale graduato nella cellula postsinaptica il neurotrasmettitore libero
viene eliminato attraverso esocitosi (può essere riciclato o distrutto da enzimi).
>Il potenziale d’azione trasporta un’info, questo potenziale è dovuto all’apertura dei canali voltaggio
dipendenti (sodio e potassio, per un valore soglia), che portano all’inversione della polarità della
membrana (in tutto l’assone) e quando ciò arriva al terminale presinaptico causa l’apertura dei
canali del calcio.
>L’acido glutammico apre un canale per il sodio, quindi il recettore per il glutammato è un canale
per il sodio; normalmente la membrana non è permeabile al sodio, ma quando il glutammato si
lega al suo recettore, quest’ultimo cambia conformazione, facendo aprire il canale e fluire il sodio.
L’acido gamma amino butirrico (GABA) fa la stessa cosa per il cloro, aprendo il canale, mentre la
dopamina apre il canale per il potassio.
Il potenziale di membrana varia a seconda dei canali ionici che si aprono (tende verso i valori
negativi in particolare, quindi verso il potenziale di equilibrio di Cl-, Na+, K+). Se la membrana
invece di depolarizzarsi si iperpolarizzasse, il campo elettrico ci sarebbe comunque, facendo
passare però una corrente iperpolarizzante. Nel caso della sinapsi chimica essa è però specifica:
se c’è una sinapsi glutammatergica, è una sinapsi eccitatoria, in quanto apre i canali per il sodio
che rende l’interno della cellula positivo; se c’è una sinapsi gabaergica che governa il canale per il
cloro, questa è inibitoria; se c’è una sinapsi dopaminergica che apre il canale per il potassio, allora
il segnale è specifico.
SPINE DENDRITICHE
Le spine dendritiche sono coinvolte nei processi di memorizzazione, sono specifiche e sono sede
di afferenze eccitatorie (glutammatergiche); non esistono spine dendritiche inibitorie. Inoltre non
hanno una morfologia fissa, bensì una che cambia costantemente nel tempo. Esse influenzano lo
stato del neurone, che, anche se di fronte a uno stesso stimolo, con la modifica della densità delle
spine risponde in modo diverso (state coding).
Queste si muovono all’interno dell’asse dendritico, cambiano forma e si adattano e possono avere
delle alterazioni congenite o acquisite
Alla nostra nascita i neuroni sono ricchi di spine dendritiche, rispetto all’età adulta fenomeno del
pruning (potatura), che rimodella i circuiti cerebrali per rafforzare le sinapsi cerebrali e per
consentirle di sopravvivere al processo di selezione che serve per mantenere le sinapsi più
efficienti, eliminando quelle inefficienti e poco utilizzate.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
CODIFICA DELLE INFORMAZIONI
L’informazione viene codificata ma non espressa nella forma del potenziale d’azione. Perché
esistano i messaggi che passano attraverso le fibre di connessione è necessario il potenziale
d’azione, ma perché quei messaggi devono essere codificati. Il codice di comunicazione neuronale
è l'insieme delle regole di corrispondenza con cui i neuroni codificano l’informazione ricevuta,
elaborata e trasmessa.
Il codice ha una componente spaziale che definisce quali cellule sono attivate in risposta ad uno
stimolo e una componente temporale che descrive le regole di corrispondenza con cui l’attività
elettrica di un neurone codifica i segnali con cui viene stimolato.
Abbiamo tre tipi di codifica, l’ensemble coding (complesso, il modo in cui i neuroni collaborano per
formare un codice), lo state encoding (quale aspetto del neurone è rilevante) e il temporal coding
(segnali interpretati sul tempo, rate coding of frequency coding).
Ensemble coding
Partiamo dall’esempio delle informazioni acustiche, che dall’orecchio, dopo alcuni step, finiscono in
un punto preciso della corteccia. Il suono non è altro che una forma di compressione delle onde
che viaggiano in un fluido (l’aria). Queste onde di compressione fanno vibrare un sistema che parte
dal timpano e finisce nella coclea (orecchio interno). La coclea è quindi un traduttore
elettromagnetico che vibrando fa muovere le ciglia di alcune cellule. Le varie frequenze che
compongono un suono fanno quindi vibrare delle zone precise della coclea e queste vibrazioni
vengono trasformate in potenziale d’azione, trasportato in seguito in un punto preciso della
corteccia e verranno lette come informazioni di tipo acustico. Tutti i potenziali d’azione che arrivano
da vie acustiche per finire in un punto specifico della corteccia sono informazioni di natura uditiva.
C’è una via specifica anche per la vista, per l’olfatto, il gusto e così via. C’è quindi una struttura
anatomica che dà un senso alle informazioni, le codifica.
Differenti fibre raggiungono differenti zone della corteccia, come le vibrisse degli animali, che sono
collegate a punti specifici della corteccia. Negli umani accade invece che ascoltando un
determinato suono/nota musicale si attivi una precisa corrispondenza nella corteccia.
State encoding
Non si osserva più la struttura anatomica, ma si guarda un unico neurone, che non è una struttura
fissa, è plastico. Il neurone davanti allo stesso input risponde in maniera differente (anche se
sempre attraverso un potenziale d’azione) a seconda delle situazioni. È importante distinguere
quello che è lo stato del neurone e quello che è il suo output (PA). Per poterlo fare è necessario
studiare il processo del calcio.
Temporal coding
Il temporal coding aiuta l’encefalo a rilevare il ritmo della sua attività elettrica. È una codifica in due
stati. C’è inoltre una relazione lineare tra la frequenza dei potenziali d’azione di una fibra nervosa
e la contrazione muscolare. La frequenza ci dà la visione quantitativa di uno stimolo: con
l'ensemble coding e lo state coding abbiamo un’idea qualitativa del messaggio, l’ampiezza del
suono viene riconosciuta appunto dal temporal coding. Tanto più l’onda sonora è ampia più la
frequenza di scarica dei potenziali d’azione sale. Possiamo codificare due stati; il potenziale
d’azione c’è o non c’è.
Se consideriamo il frequency coding possiamo codificare tre stati: uno in cui il PA arriva ad uno
stato normale, uno in cui i PA sono in una frequenza superiore e uno in cui i PA sono in una
frequenza molto inferiore. A seconda della frequenza abbiamo un’informazione di tipo quantitativo
che va a sommarsi con un’informazione di tipo qualitativo, i neuroni quindi comunicano in FM
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
(modulazione di frequenza). Il firing del potenziale d’azione è regolato dalle afferenze che
arrivano dagli altri neuroni, dalle info che arrivano alle sinapsi elettriche o chimiche che
provengono dal mondo circostante o da altre zone cerebrali. A seconda delle condizioni il firing può
cambiare, anche in risposta ad uno stesso stimolo.
RUOLO STRUTTURA ANATOMICA GESTIONE DELLE INFO
La struttura cerebrale è composta da sei strati e le info giungono nel quarto. Ognuno di questi
strati ha inoltre un’organizzazione somatotopica, esiste quindi una correlazione geografica tra un
punto della corteccia e uno dell’organismo.
Il cervello presenta 4 lobi superficiali in
ogni emisfero. I lobi prendono il nome
dall’osso cranico sotto il quale si
trovano. Esistono due lobi profondi: il
lobo limbico e il lobo dell’insula. La
corteccia cerebrale è specializzata in
aree sensoriali per la percezione, aree
motorie che dirigono il movimento e
aree di associazione che integrano
l’informazione. A queste due zone in
particolare corrispondono l’homunculus
sensitivo e l’homunculus motorio.
Ad aree adiacenti dello spazio recettivo
corrispondono gruppi di neuroni
adiacenti in un’area corticale, per la
corteccia motoria in particolare gruppi di neuroni adiacenti governano muscoli anatomicamente
adiacenti.
L’homunculus, che sia sensitivo o motorio, è dismorfico, perché la rappresentazione è in termini di
superficie della corteccia dedicata a quella particolare funzione. L’homunculus motorio ha mani e
lingua enormi, soprattutto se confrontati al piede, in quanto a livello di queste zone abbiamo un
controllo motorio (soprattutto nelle dita) molto più fine e di conseguenza abbiamo bisogno di tanta
corteccia per controllare il movimento, il braccio invece è rappresentato molto piccolo in quanto non
serve per compiere movimenti fini. L’homunculus sensitivo invece ha mani enormi, in quanto la
mano è molto sensibile e riesce a capire cosa tocca senza vederlo, abbiamo sensibilità
millimetrica, come nella lingua. Anche le gengive hanno una sensibilità loro, però non avendo
muscoli sono rappresentate solo nel secondo homunculus.
Alterazioni alla struttura encefalica (malformazioni congenite):
-lissencefalia : encefalo liscio, le circonvoluzioni cerebrali sono assenti
-pachigiria: circonvoluzioni cerebrali scoordinate
-schizencefalia: zone cerebrali disorganizzate
TAC - TOMOGRAFIA ASSIALE COMPUTERIZZATA
Si basa sull'emissione di raggi x (fotoni ad alta energia) che vengono emessi da un lato
dell'anello e assorbiti dal lato opposto. Il paziente viene messo al centro dell'anello, la struttura
lavora sul tessuto biologico dell'encefalo e i raggi x registrano diversamente l'osso, il liquido
cerebrospinale e vari tessuti (sostanza grigia e sostanza bianca). In base a quanti raggi sono
stati assorbiti si va a ricostruire l'immagine di ciascuna "fetta" dell'encefalo.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Questo permette di andare a ricostruire delle mappe colorimetriche che identificano le diverse
strutture cerebrali.
↳i raggi vengono assorbiti più o meno a seconda della presenza del calcio:
↪ quindi la teca cranica che contiene molto calcio viene visualizzata come molto chiara,
mentre la sostanza grigia, che si trova immediatamente dopo la teca cranica viene visualizzata
più bianca rispetto alla sostanza bianca (immediatamente dopo).
IL fatto che la sostanza grigia venga visualizzata più bianca rispetto alla sostanza bianca è
dovuto al fatto che il sangue (in maggiori quantità nella sostanza grigia, più vascolarizzata)
contiene calcio. La sostanza bianca è meno vascolarizzata, quindi meno bianca.
Successivamente quello più scuro è il liquor, composto prevalentemente da acqua.
STIMOLAZIONE MAGNETICA TRANSCRANICA (TMS)
Tecnica di stimolazione corticale non invasiva, sicura e non dolorosa, induzione elettromagnetica
regolata dalla legge di Faraday. Il campo magnetico è capace di indurre un campo elettrico, e
quindi di conseguenza una corrente elettrica in un tessuto eccitabile.
Lo stimolatore consiste di due parti distinte:
•un generatore e accumulatore di corrente,
•una bobina di spire di rame (coil) in cui il passaggio della corrente induce un campo magnetico ad
alta intensità e di breve durata, con linee di flusso perpendicolari al piano del coil.
>Il campo magnetico generato nel coil attraversa la cute e il cranio e raggiunge facilmente il
tessuto cerebrale sottostante, inducendo un campo elettrico che ha la capacità di depolarizzare
la membrana degli elementi cerebrali eccitabili.
Profondità: pochi cm, non può raggiungere strutture profonde.
>La TMS è stata sviluppata per scopi clinici (valutare l’integrità della via corticospinale), ma poi
è divenuto un utile strumento di ricerca grazie alla capacità di alterare la normale attività di una
regione specifica della corteccia . In Modo simile, degli studi basati su lesioni, le conseguenze
comportamentali prodotte dalla stimolazione possono essere utili per far luce sul funzionamento
normale dell’area trattata.
tDCS STIMOLAZIONE TRANSCRANICA A CORRENTE CONTINUA
La tDCS è una tecnica di facile applicazione che permette di stimolare diverse parti del cervello
in modo non invasivo, indolore e senza effetti collaterali significativi. La stimolazione consiste in
una corrente elettrica continua a bassa intensità (1-2 mA), non percepita dalla persona, per
periodi prolungati di tempo (5-30 minuti). La corrente viene applicata allo scalpo tramite una
coppia di elettrodi (uno eccitatorio, anodo, e uno inibitorio, catodo) e permette così di
influenzare l’eccitabilità corticale, e quindi l’attività neuronale, della regione posta sotto gli
elettrodi stessi.
DEEP BRAIN STIMULATION (DBS)
L’intervento neurochirurgico usato per ristabilire la normale funzionalità. La stimolazione
cerebrale profonda (DBS dall'acronimo inglese Deep Brain Stimulation) è un trattamento
chirurgico volto a ridurre i sintomi motori debilitanti caratteristici dei disturbi del movimento
come il Parkinson, la distonia e il tremore essenziale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Consiste nell'inserimento di un elettrodo in una zona specifica del cervello, collegato ad un
neurostimolatore posto nella zona superiore del torace o nell'addome. Il neurostimolatore invia
impulsi elettrici al cervello, finalizzati a ridurre i sintomi della patologia specifica.
L’elettrocatetere è impiantato nel nucleo subtalamico nel caso del morbo di Parkinson, nel
nucleo ventrale intermedio se usato per alleviare il tremore essenziale e nel nucleo del globo
pallido per le distonie: stimolando i vari nuclei si bloccano i segnali che provocano i sintomi
motori disabilitanti delle malattie.
EEG – ELETTROENCEFALOGRAMMA
L’EEG misura un derivato dell’attività elettrica, che dipende dai potenziali post sinaptici e riflette
l’attività dei gruppi neuronali. La registrazione di questo derivato dell’attività elettrica si svolge con
gli elettrodi. Il potenziale elettrico che si registra sulla superficie del cranio proviene dai neuroni, ma
non è il potenziale d’azione: dipende appunto dai potenziali post sinaptici che possono essere
eccitatori o inibitori. In entrambi i casi essi vanno a formare un dipolo (una parte carica
positivamente e l’altra carica negativamente) che va a creare un campo elettrico. Si tratta
dell’attività di gruppi di neuroni, in particolare di quelli piramidali, che sono allineati in fila sulla
corteccia (divisa a sua volta in lobi), poiché la corteccia è la parte più vicina e che meglio può
interagire con l’elettrodo. Il dipolo può essere letto in diversi modi a seconda della parte che
osserviamo: possiamo vederne la parte positiva o negativa, oppure nessuna delle due.
>Ritmi dell’attività elettrica → registrando il campo elettrico dell’encefalo si scopre che esso ha
natura oscillatoria, perché i neuroni eccitatori attivano gli inibitori a livello corticale (i neuroni
eccitatori sono legati tra loro e il firing si attiva grazie agli eccitatori e si spegne grazie agli
inibitori).
>Natura oscillatoria →frequenze lente, più ampie, oppure più frequenti ma meno ampie. La
frequenza degli stati oscillatori varia a seconda dello stato d’animo o di determinate patologie.
>I tracciati EEG ottenuti in una serie di prove vengono mediati tra loro sincronizzando le
registrazioni rispetto ad un evento esterno, es: la comparsa di uno stimolo o di una risposta. Gli
ERP sono risposte molto piccole, nascoste nel flusso dell’EEG corrente. Calcolando la media
delle tracce EEG si possono estrarre le risposte, che riflettono l’attività neurale specificatamente
collegata a un particolare evento sensoriale, motorio, cognitivo.
MIDOLLO SPINALE
E’ la principale via in cui le informazioni corrono bidirezionalmente tra encefalo e periferia. I
neuroni del midollo spinale indirizzano le informazioni dai recettori periferici all'encefalo e gli
“ordini” dall’encefalo agli effettori (muscoli e ghiandole). Inoltre contiene le reti neurali
responsabili della locomozione.
IPOTALAMO
È un'area che si affaccia sulla superficie inferiore dell'encefalo. è collocato sotto il talamo e
l’ipofisi; prende i contatti con: encefalo/ tronco dell'encefalo/ sistema limbico/ midollo spinale
Funzioni:
-Media gli stimoli per la sopravvivenza
-Responsabilità di elaborazione delle emozioni
-Controllo del sistema nervoso autonomo
-Agisce sul sistema endocrino
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
LA VISIONE
L’occhio è l’organo deputato alla visione e le sue strutture principali sono la cavità orbitaria, il
bulbo oculare e le vie ottiche. La visione è il risultato di un'interpretazione della realtà fisica
elaborata dal cervello; a volte questa interpretazione può essere errata come nel caso delle
illusioni ottiche.
ANATOMIA DELL’OCCHIO
La pupilla è un’apertura che permette alla luce di entrare nell’occhio e raggiungere la retina. Le
dimensioni della pupilla sono controllate da un muscolo circolare: l’iride (colore dell’occhio),
contiene due muscoli uno che quando si contrae restringe la pupilla, l’altro la ingrandisce. La
pupilla e l’iride sono ricoperte dalla superficie esterna vitrea e trasparente dell’occhio: la cornea.
La cornea prosegue nella sclera, «il bianco dell’occhio», la parte dura del globo oculare. Nella
sclera sono inseriti tre paia di muscoli che fanno muovere il globo oculare nell’orbita ossea.
Questi non sono visibili perché giacciono sotto la congiuntiva, una membrana ripiegata che
nasce dalla parte interna delle palpebre e si congiunge alla sclera. Il nervo ottico, che trasporta
gli assoni della retina, fuoriesce dal retro dell’occhio. Passa attraverso l’orbita e raggiunge il
cervello alla sua base, in prossimità dell’ipofisi.
CRISTALLINO E FORMAZIONE DELLE IMMAGINI SULLA RETINA
Dietro la pupilla si trova il cristallino, struttura avascolare, trasparente ed elastica, sospesa da
fibre che costituiscono la zonula di Zinn (noto anche come zonula ciliare o Apparato
sospensore del cristallino). È un sistema di fibre tese fra il cristallino e il corpo ciliare. Il corpo
ciliare è un anello di tessuto che circonda il cristallino, consiste di una componente muscolare
importante per l’aggiustamento del potere di rifrazione del cristallino e di una componente
vascolare (processi ciliari) che produce un fluido che riempie la parte frontale dell’occhio.
Oltrepassata la cornea e prima di colpire la retina i raggi luminosi passano attraverso due
diversi ambienti liquidi.
La cavità posteriore è situata dietro il cristallino, è occupata dal corpo vitreo. È costituita da una
sostanza incolore, trasparente, gelatinosa e vischiosa con diverse funzioni:
· Statica: consente grazie alla propria pressione il mantenimento della forma e del volume
del globo oculare
· Trofica: interviene nel nutrimento delle strutture avascolari (cornea, cristallino) fornendo
ossigeno e glucosio e allontanando impurità (mosche volanti)
La cavità anteriore è lo spazio tra cristallino e cornea dove si trova l’umore acqueo, liquido
acquoso trasparente che rifornisce di sostanze nutritive queste due strutture.
L’umor acqueo è prodotto dai processi ciliari nella camera posteriore (posta tra cristallino e
iride) e fluisce nella camera anteriore attraverso la pupilla.
Per una normale visione è necessario che i mezzi ottici dell’occhio siano trasparenti e cornea e
cristallino sono due esempi di specializzazione tissutale formidabili per questo scopo. Oltre a
garantire un’efficiente trasmissione della luce, la funzione principale delle componenti
dell’occhio è quella di ottenere la formazione di un’immagine a fuoco sulla superficie della
retina.
La cornea e il cristallino sono i maggiori responsabili della deviazione (rifrazione) dei raggi
luminosi sulla retina. Di conseguenza si ha la formazione di un’immagine a fuoco sulla retina →
cornea (indice rifrazione simile a quello dell’acqua).
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Il cristallino ha un potere di rifrazione molto inferiore rispetto alla cornea però è regolabile e
rende possibile la messa a fuoco nitida di oggetti posti a varia distanza.
Le modificazioni dinamiche del potere di rifrazione del cristallino prendono il nome di
accomodazione: elasticità intrinseca/fibre zonulari→ muscolo ciliare.
Visione oggetti distanti: il cristallino si appiattisce e diviene relativamente sottile, assumendo il
valore minimo del suo potere di rifrazione → Oggetti distanti: la forza delle fibre zonulari
dell’elasticità del cristallino che diventa piatto.
Visione oggetti vicini: il cristallino diviene più spesso e arrotondato, assumendo il massimo del
suo potere di rifrazione → Oggetti vicini: la forza delle fibre zonulari dell’elasticità del cristallino
che prevale e aumenta la curvatura.
CAMBIAMENTI DOVUTI AL MUSCOLO CILIARE CHE CIRCONDA IL CRISTALLINO
Il cristallino è mantenuto in sede da fibre radiali di connettivo: fibre zonulari.
La forma del cristallino è dovuta a due forze opposte: la sua elasticità che tende a mantenere la
forma tonda e la tensione delle fibre zonulari.
Il cristallino è legato al muscolo ciliare dalle fibre anelestiche, dette collettivamente zonula.
Quando il muscolo ciliare è rilassato, la zonula tira sul cristallino e lo mantiene appiattito.
Quando è contratto diminuisce la tensione della zonula e il cristallino, che è elastico, assume
una forma più rotonda.
Quando i raggi luminosi paralleli passano attraverso il cristallino appiattito, il punto focale cade
sulla retina. Nel caso di oggetti vicini, i raggi luminosi non sono più paralleli; il cristallino e la sua
distanza focale non si sono modificati, ma l’oggetto è visto fuori fuoco perché i raggi luminosi
non sono a fuoco sulla retina.
A differenza del cristallino appiattito, il cristallino arrotondato ha una distanza focale minore che
è adatto alla visione da vicino; quindi per mantenere a fuoco un oggetto che si avvicina, il
cristallino si arrotonda.
LA RETINA VISTA CON L’OFTALMOSCOPIO
La fovea è la zona retinica dove la visione è più acuta e corrisponde al centro dello sguardo che
rivolgiamo agli oggetti che attirano la nostra attenzione. È anche la zona dove i fotorecettori,
delle cellule bipolari e gangliari è massima. Al centro si trova la foveola, dove gli strati cellulari
sono spostati lateralmente per ridurre l’ulteriore offuscamento dovuto alla rifrazione della luce.
STRUTTURA DELLA RETINA
I fotorecettori sono cellule nervose che si trovano sulla retina. Sono recettori sensoriali sensibili
ai fotoni e svolgono un'importante funzione di trasduzione, cioè sono in grado di trasformare la
luce che arriva sul fondo dell'occhio in un segnale elettrico da trasmettere al cervello mediante il
nervo ottico. I fotorecettori della retina sono distinti in coni e bastoncelli.
Oltre ai fotorecettori si trovano anche le cellule bipolari, orizzontali, amacrine e gangliari.
I coni e i bastoncelli sono i recettori della luce e generano i segnali elettrici che, instradati sulle
vie ottiche raggiungono l’area della visione della corteccia. I coni e i bastoncelli contengono
molecole fotosensibili che assorbono la radiazione luminosa e producono segnale elettrico.
Coni) sono impiegati per la visione diurna e il riconoscimento dei colori. sono concentrati
principalmente nella fovea retinica e sono determinanti per la qualità visiva.
-3 tipi: corrispondenti al colore rosso, verde e blu
Bastoncelli) più numerosi, hanno un elevata sensibilità alla luce e consentono la visione anche
con scarsa illuminazione. (visione notturna)
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Gli strati della retina:
1. Epitelio pigmentato
2. Segmenti esterni dei fotorecettori: contiene gli elementi fotosensibili della retina. I segmenti
esterni sono immersi nell’epitelio pigmentato
3. Strato nucleare esterno: contiene i corpi cellulari dei fotorecettori
4. Strato plessiforme esterno: contiene i contatti sinaptici tra i fotorecettori e le cellule bipolari
e orizzontali
5. Strato nucleare interno: contiene i corpi cellulari delle cellule bipolari, orizzontali ed
amacrine
6. Strato plessiforme interno: contiene i contatti sinaptici fra le cellule bipolari, amacrine e
gangliari
7. Strato delle cellule gangliari: contiene i corpi cellulari delle cellule gangliari
8. Strato delle fibre nervose
EPITELIO PIGMENTATO
A prima vista, la disposizione spaziale degli strati retinici sembra contro intuitiva, poiché i raggi
di luce devono passare attraverso vari elementi della retina non sensibili alla luce, nonché
attraverso la coroide della retina (vascolarizzazione della retina) che si ramifica ampiamente
sulla sua superficie interna prima di raggiungere i segmenti esterni dei fotorecettori, in cui i
fotoni vengono assorbiti.
La ragione di questa curiosa caratteristica dell'organizzazione retinica risiede nella relazione
speciale che esiste tra i segmenti esterni dei fotorecettori, l'epitelio del pigmento e la coroide
sottostante.
I capillari nella coroide sottostante all'epitelio pigmentato sono la fonte primaria di nutrimento
per i fotorecettori retinici. Queste considerazioni funzionali spiegano presumibilmente il motivo
per cui bastoncelli e coni si trovano nello strato più esterno piuttosto che più interno della
retina.
La retina è in stretto rapporto con l’epitelio pigmentato, le cui cellule contengono grandi
quantità di melanina, un pigmento nero che assorbe la luce che non è stata trattenuta dalla
retina e impedisce che possa essere riflessa su altre parti della retina stessa, con
conseguente deterioramento delle immagini visive.
I segmenti esterni dei fotorecettori contengono dischi membranosi che ospitano il
fotopigmento sensibile alla luce e altre proteine coinvolte nel processo di trasduzione. Questi
dischi si formano vicino al segmento interno del fotorecettore e si spostano verso la punta del
segmento esterno.
L'epitelio pigmentato svolge un ruolo essenziale nel fagocitare le estremità del segmento
esterno facilitandone il ricambio; questo non è un compito da poco, poiché tutti i dischi nei
segmenti esterni vengono sostituiti ogni 12 giorni. Inoltre, l'epitelio pigmentato contiene i
macchinari biochimici necessari per rigenerare le molecole di fotopigmento dopo che sono
state esposte alla luce.
La durata della vita del fotorecettore è visualizzata tracciando lo spostamento di amminoacidi
marcati radioattivamente che, dopo essere stati iniettati nel segmento interno, sono stati
incorporati nei dischi. I dischi così marcati migrano dalla porzione interna a quella esterna del
segmento esterno in un periodo di 12 giorni.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
I dischi che hanno terminato il loro ciclo biologico sono eliminati dal segmento esterno e
fagocitati. Il fotopigmento contenuto nei dischi entra nell’epitelio pigmentato dove verrà
riciclato biochimicamente e reinserito nei dischi neoformati del fotorecettore.
RETINITE PIGMENTOSA
I capillari nella coroide sottostante all'epitelio pigmentato sono la fonte primaria di nutrimento
per i fotorecettori retinici. Queste considerazioni funzionali spiegano perché le interruzioni
nelle normali relazioni tra l'epitelio del pigmento e i fotorecettori retinici come quelli che si
verificano nella retinite pigmentosa hanno gravi conseguenze per la vista.
FOTORECETTORI A CONFRONTO
Sia i coni che i bastoncelli sono formati da due porzioni: segmento interno e segmento
esterno. Il segmento esterno contiene invaginazioni della membrana che formano strati simili
a dischi membranosi che contengono il fotopigmento. Il segmento interno contiene il nucleo
cellulare e i vari organuli cellulari. All’estremità del segmento interno si trova il terminale
sinaptico ricco di vescicole sinaptiche contenenti il neurotrasmettitore.
SEGMENTO ESTERNO A FORMA DI SEGMENTO ESTERNO A FORMA DI
BASTONCELLO CONO
120.000.000 PER OCCHIO, DISTRUBUITI 6.400.000 PER OCCHIO, LOCALIZZATI
IN TUTTA LA RETINA (VISIONE SOPRATTUTTO NELLA FOVEA (VISIONE
PERIFERICA) CENTRALE)
BUONA SENSIBILITÀ POVERA SENSIBILITÀ
1 TIPO → VISIONE MONOCROMATICA 3 TIPI (R, G, B) → VISIONE A COLORI
MOLTI BASRONCELLI SI CONNETTONO OGNI CONO È CONNESSO AD UNA
AD UNA CELLULA BIPOLARE → SCARSA SOLA CELLULA BIPOLARE → BUONA
ACUITÀ = POVERA RISOLUZIONE ACUITÀ = BUONA RISOLUZIONE
VISIONE
Nelle cellule bipolari non si ha ancora potenziale d'azione e l'impulso elettrico, iperpolarizzante
o depolarizzante, generato dai canali sensibili al glutammato, viene condotto
elettrotonicamente. Se una cellula bipolare viene depolarizzata, essa secernerà
neurotrasmettitori eccitatori alle cellule gangliari, poste più internamente, nelle quali si genererà
il potenziale d'azione.
Le cellule orizzontali ricevono afferenze dai fotorecettori e proiettano i loro neuriti lateralmente
per influenzare le circostanti cellule bipolari e i fotorecettori.
Diverse cellule amacrine ricevono afferenze dalle cellule bipolari e proiettano lateralmente per
influenzare le circostanti cellule gangliari, le cellule bipolari e le cellule amacrine.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Le cellule gangliari sono gli unici neuroni retinici che generano potenziali d’azione, essenziali
per trasmettere le informazioni fuori dall’occhio. I loro assoni convergono tutti verso il punto
cieco della retina, là dove si uniscono per formare il nervo ottico.
L’assorbimento della luce da parte del fotopigmento nel segmento esterno dei fotorecettori
avvia una cascata di eventi che modifica il potenziale di membrana del recettore, quindi la
quantità di neurotrasmettitore rilasciata dal fotorecettore fa sinapsi sulle cellule a contatto.
Nello strato plessiforme esterno, tra i terminali dei fotorecettori e le cellule bipolari (e le cellule
orizzontali) si formano delle sinapsi; più specificamente, i corpi cellulari dei fotorecettori formano
lo strato nucleare esterno, mentre i corpi cellulari delle cellule bipolari si trovano nello strato
nucleare interno. I brevi processi assonali delle cellule bipolari stabiliscono contatti sinaptici a
loro volta sui processi dendritici delle cellule gangliari nello strato plessiforme interno.
Gli assoni molto più grandi delle cellule gangliari formano il nervo ottico e trasportano
informazioni sulla stimolazione retinica verso le altre parti del sistema nervoso centrale.
FOTOTRASDUZIONE
è quel processo tramite il quale la luce viene convertita da segnali nervosi. Avviene tramite
delle molecole chiamate fotopigmenti → si trovano all’interno dei fotorecettori che occupano la
parte posteriore della retina.
-da un punto di vista anatomico un bastoncello possiede un segmento esterno e uno interno.
Nel segmento esterno troviamo dei piccoli dischi che si originano dalla membrana plasmatica e
contengono i fotopigmenti.
>processo: quando c’è una condizione di buio vi è un continuo rilascio di glutammato;
quando arriva la luce il rilascio di glutammato da parte del bastoncello finisce. Al buio vi
sono dei canali cationici che lasciano entrare all’interno della cellula Na e Ca. Questi canali
al buio sono canali dipendenti dalla molecola cGMP. La concentrazione di cGMP al buio è
alta, si lega ai canali i quali si aprono lasciando l’accesso al Na e Ca, cosi che al buio c’è
una continua depolarizzazione. Quando arriva la luce succede che la luce contatta i
fotopigmenti che tramite diverse reazioni vanno a ridurre la concentrazione di cGMP.
Questo abbassamento fa chiudere i canali cationici, non entrano più Na e Ca, non c’è più
depolarizzazione e non c’è più rilascio di neurotrasmettitore.
DAL PUNTO DI VISTA MOLECOLARE → INTERNO DEL DISCO DEL BASTONCELLO
nella membrana è incastonata una proteina di transmembrana ovvero il fotopigmento che nel
caso dei bastoncelli si chiama rodopsina:
-opsina = proteina/ 11-cis retinale = molecola organica.
la luce colpisce la rodopsina che viene assorbita dall’11- cis retinale, che grazie alla luce
subisce un combiamento strutturale, si isomizza diventando retinale tutto trans. Tutto ciò
provoca un cambiamento conformazionale anche dell’ospina e così tutta la molecola di
rodopsina grazie alla luce si converte in metarodopsina = molecola chiave nel processo di
fototrasduzione.
La metarodopsina va a contattare una proteina “g” che si chiama trasducina. Appena si forma il
legame tra la metarodopsina e la trasducina, la subunità alpha della proteina g, si stacca e
contatta un enzima di membrana = la fosfodiesterasi → catalizza la reazione che converte il
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
cGMP in GMP. Grazie alla PDE il cGMP viene convertito in GMP e quindi non ci sono più
molecole che possono legarsi ai canali per farli stare aperti.
-di conseguenza all’interno della cellula avremo una iperpolarizzazione perché canali cationici
del Na e del Ca sono chiusi, prendono il dominio i canali del K. Avremo un afflusso di K, esce
dalla cellula e la rende più negativa.
COME LA RODOPSINA RISPONDE AD UN ALTRO FOTONE
la rodopsina è nello stato di metarodopsina. Bisogna bloccare le reazioni avvenute
precedentemente, lo si fa andando a fosforilare (attaccare un gruppo fosfato alla metarodopsina
per il tramite della rodopsina chinasi e dopodiché alla metarodopsina si va ad attaccare un’altra
proteina = arrestina, la quale arresta il funzionamento della metarodopsina spezzandola.
Appaiono l’opsina e il retinale tutto trans il quale viene passato all’epitelio pigmentato che si
trova sul retro della retina. Viene rinconvertito in 11 cis, si unisce all’opsina e si riottiene la
rodopsina e il ciclo potrà ripartire da capo.
IL CICLO DEL RETINOLO
Il retinolo tutto-trans (vitamina A) è il precursore del retinale 11- cis. I suoi precursori come la
vitamina A non vengono sintetizzati dall’uomo e devono essere assunti con la dieta. La carenza
di VIT-A porta alla cecità notturna.
Dopo la fotoisomerizzazione il retinale tutto-trans è convertito in retinolo tutto-trans ed è
trasportato ad opera della proteina chaperon IRBP (Interphotoreceptor retinoid binding protein)
nell’epitelio pigmentato. Qui, in un processo a più tappe è riconvertito a retinale 11-cis e
trasferito nuovamente (sempre tramite la IRBP) al segmento esterno del fotorecettore dove si
ricombina con l’opsina.
AMPLIFICAZIONE DEL SEGNALE
Una delle caratteristiche importanti di questa complessa cascata biochimica avviata dalla
cattura di fotoni è che fornisce un'enorme amplificazione del segnale.
È stato stimato che: una singola molecola di rodopsina attivata dalla luce può attivare 800
molecole di trasducina, circa l'8% delle molecole di trasducina sulla superficie del disco.
Sebbene ogni molecola di trasducina attivi solo 1 molecola di fosfodiesterasi, ognuna di queste
è a sua volta in grado di catalizzare la rottura di ben 6 molecole di cGMP. Di conseguenza,
l'assorbimento di un singolo fotone da parte di una molecola di rodopsina provoca la chiusura di
circa 200 canali ionici (circa il 2% del numero di canali in ciascun bastoncello). La chiusura di
questo numero di canali provoca una variazione netta del potenziale di membrana di circa 1
mV.
È importante sottolineare che questa cascata di amplificazione viene interrotta una volta
completato il ciclo visivo e le molecole tornano al loro stato inattivato: la rodopsina attivata viene
fosforilata da una rodopsina chinasi e questa modifica consente a un'altra proteina, chiamata
arrestina, di unirsi alla rodopsina. Il complesso rodopsina-arrestina non può attivare la
trasducina, quindi la cascata di fosforilazione viene interrotta.
RIASSUMENDO
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
FOTORECETTORE AL BUIO
In condizioni di buio la concentrazione intracellulare di GMP ciclico è elevata all’interno del
segmento esterno del fotorecettore.
L’elevata concentrazione di GMP ciclico mantiene aperti i canali CNGC (canali cationici attivati
da nucleotidi ciclici). Essi generano una corrente entrante depolarizzante, trasportata
principalmente da Na+ , ma anche Ca2+ , dette corrente di buio. Questa corrente è
controbilanciata da una corrente uscente che fluisce attraverso la membrana del segmento
interno, dovuta a canali per il K+ (attivati dalla depolarizzazione della membrana).
L’effetto complessivo è che il fotorecettore si trova depolarizzato ad un «potenziale di buio» di
-40 mV. A livello della terminazione sinaptica, la depolarizzazione della membrana del
fotorecettore induce l’apertura dei canali del Ca2+ . L’ingresso del Ca2+ attiva l’esocitosi al buio
del neurotrasmettitore (glutammato) che induce la formazione di un potenziale graduato nella
cellula bipolare.
FOTORECETTORE ALLA LUCE
Quando il fotorecettore è esposto alla luce, il retinale cambia conformazione e si dissocia
dall’opsina che cambia conformazione in metarodopsina II. Questa attiva la trasducina che a
sua volta attiva la fosfodiesterasi che catalizza la scissione del GMP ciclico. La diminuzione
della concentrazione di GMP ciclico nel segmento esterno provoca la chiusura dei canali
CNGC. Di conseguenza la membrana si iperpolarizza determinando la chiusura dei canali del
Ca2+ nel segmento interno e conseguente diminuzione della liberazione di neurotrasmettitore.
IL RUOLO DEL Ca 2+
Nel segmento esterno, il calcio
inibisce l’attività della guanilato
ciclasi e della rodopsina chinasi,
oltre a ridurre l’affinità dei canali
cGMP-dipendenti per il cGMP. La
chiusura, indotta dalla luce, dei
canali della membrana del
segmento esterno porta a una
riduzione della concentrazione di
ioni Ca2+ e a una riduzione Ca2+
dipendente di questi elementi della
cascata. Di conseguenza, si riduce
la fotosensibilità del recettore alla
luce.
SPECIALIZZAZIONI FUNZIONALI DEI CONI E DEI BASTONCELLI
Differenze strutturali e funzionali tra bastoncelli e coni. Sebbene generalmente simili nella
struttura, bastoncelli e coni differiscono per dimensioni e forma, così come per la disposizione
dei dischi membranosi nei loro segmenti esterni e per le caratteristiche delle loro connessioni
sinaptiche.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Queste proprietà riflettono il fatto che i sistemi dei bastoncelli e dei coni sono specializzati per
diversi aspetti della visione.
Il sistema dei bastoncelli ha una risoluzione spaziale molto bassa ma è estremamente sensibile
alla luce; è quindi specializzato per la sensibilità a spese della risoluzione.
Il sistema dei coni ha una risoluzione spaziale molto elevata, ma è relativamente insensibile alla
luce; è quindi specializzato per l'acuità a spese della sensibilità. Le proprietà del sistema a cono
consentono all'uomo e a molti altri animali di vedere il colore.
Ai livelli più bassi di illuminazione, vengono attivate solo i bastoncelli. I coni iniziano a
contribuire alla percezione circa al livello della luce delle stelle e sono gli unici recettori che
funzionano in condizioni relativamente luminose.
Bastoncelli → rilevatori di luce
Ogni cellula bipolare dei bastoncelli riceve segnali sinaptici da 15-30 bastoncelli. Più a valle in
questa stessa via si ha un ulteriore incremento del livello di convergenza, in quanto molte
cellule bipolari dei bastoncelli convergono su una data cellula amacrina.
Coni → massima acuità
Al centro della fovea, invece, ogni cono prende contatto con una singola cellula bipolare e
questa, a sua volta, trasmette il segnale ad una singola cellula gangliare, che ha un ruolo
dominante nella visione centrale
DISTRIBUZIONE DEI FOTORECETTORI NELLA RETINA UMANA
L’aumento dei coni nella foveola si accompagna a un evidente riduzione del diametro del
segmento esterno. Sezione trasversale della fovea. Gli strati cellulari sono spostati di lato in
modo che la luce subisca la minima dispersione prima che i fotoni colpiscano i segmenti esterni
dei coni.
La densità dei coni presenta un picco in corrispondenza del centro della fovea (foveola), mentre
è basso in tutto il resto della retina.
La densità dei bastoncelli è elevata nella maggior parte della retina, ma crolla nella fovea; nella
foveola i bastoncelli sono completamente assenti.
I CONI E LA VISIONE DEI COLORI
La luce è definita come radiazione elettromagnetica con lunghezze d'onda comprese tra ~ 400
e 700 nm. L'assorbanza è definita come il log del rapporto tra l'intensità della luce incidente e
l'intensità della luce trasmessa.
Utilizzando una nuova tecnica nota come "ottica adattativa" e trucchi di adattamento alla luce è
possibile mappare con alta precisione la distribuzione dei diversi tipi di coni nella retina. Sono
stati usati falsi colori per identificare i coni per le lunghezza d'onda corte (blu), medie (verde) e
lunghe (rosso).
LA VISIONE TRICROMATICA
· Protanopia (primo difetto del colore): confondono il rosso e il verde. Esse vedono il mondo
in sfumature di giallo e blu; sia il rosso che il verde appaiono giallastri ai loro occhi. La loro
acuità visiva è normale. Sono privi dei coni sensibili alle lunghezze d’onda lunghe (rossi).
· Deuteranopia (secondo difetto del colore): confondono il rosso e il verde e la loro acuità
visiva è normale. In questo caso sono privi dei coni sensibili alle lunghezze d’onda medie
(verdi).
· Tritanopia (terzo difetto del colore): vedono il mondo in rosso e verde, l’azzurro è verde
chiaro e il giallo appare rosa. Sono privi dei coni sensibili alle lunghezze d’onda corte (blu).
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
CAMPI RECETTIVI ON/OFF DELLE CELLULE GANGLIARI - INIBIZIONE LATERALE
organizzazione della retina: i coni fanno sinapsi sulle cellule bipolari che fanno di conseguenza
sinapsi sulle gangliari = connessione verticali
tuttavia all’interno della retina ci sono connessioni orizzontali che sono mediate da degli
interneuroni inibitori come le cellule orizzontali e le cellule amacrine.
cellule orizzontali = vanno a collegare i coni ad altri coni portando un effetto inibitorio
°il campo recettivo di una cellula gangliare è dato dalla somma dei campi recettivi di tutti i coni
che convergono su di essa
>i campi recettivi delle cellule gangliari hanno un centro e una periferia. Alcuni coni formano il
centro del campo recettivo, altri la periferia.
coni verticali → formano il centro → tutti quei coni che fanno sinapsi sulle bipolari, che fanno
sinapsi sulle gangliari.
coni che formano la periferia → coni coinvolti in una connessione orizzontale
- il campo recettivo di una gangliare può essere al centro ON e alla periferia OFF
oppure centro off e periferia on
↓
centro on: risponde bene alla luce > on = luce / off = buio
periferia off: risponde belle al buio
perché c’è on e off => perché esistono due tipi di cellule bipolari → bip. on / bip. off
cambia il tipo di recettore del glutammato.
- le cellule ON avranno un recettore metabotropico che in risposta al glutammato
vanno a chiudere i canali cationici causando iperpolarizzazione → il glutammato
sulle cell. on ha un effetto inibitorio
- le cellule OFF hanno canali del glutammato ionotropici → il glutammato ha un
effetto eccitatorio
i coni rilasciano glut. quando si trovano al buio → quando arriva la luce, il rilascio di glut.
diminuisce. bip. on → glut. effetto inibitorio quindi se ce n’è meno non avrò una inibizione ma
darò spazio ad una depolarizzazione e in modo graduale rilascia neurotrasmettitore che va
sulla cellula gangliare.
periferia off → colpita del buio → il cono rilascia più glut → effetto eccitatorio sulle cellule
orizzontali → queste inibiscono i coni e le cellule bipolari; se un cono viene inibito rilascia
meno glutammato → la bipolare si depolarizza di più.
i campi recettivi delle cellule gangliari sono fatti per rispondere ai contrasti
se vi è luce nel centro e buio in periferia rispondono bene perché c’è contrasto.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
CIRCUITI ALL’ORIGINE DELLA FUNZIONE ANTAGONISTICA DELLA PERIFERIA DEL
CAMPO RICETTIVO DI UNA CELL GANGLIARE CENTRO-ON DELLA RETINA
Il risultato di questo circuito è la soppressione dei segnali provenienti dalla periferia del campo
ricettivo di una cellula gangliare centrale.
Risposte dei vari tipi cellulari alla presentazione di uno stimolo luminoso al centro del campo
ricettivo seguito dall‘illuminazione della periferia. La stimolazione luminosa della periferia
causa l'iperpolarizzazione delle cellule orizzontali e alla riduzione del rilascio del trasmettitore
inibitorio (GABA) sui terminali del fotorecettore.
L'effetto netto è di depolarizzare il terminale del cono centrale, compensando gran parte
dell'iperpolarizzazione indotta dalla cascata di trasduzione nel segmento esterno del cono
centrale.
LE VIE CENTRALI
Le informazioni visive fornite dalla retina danno origine a interazioni tra diverse aree
dell’encefalo che culminano nella percezione cosciente della scena visiva e allo stesso tempo,
stimolano i riflessi più comuni come quelli relativi all’aggiustamento della dimensione della
pupilla, all’orientamento degli occhi verso gli oggetti di interesse e alla regolazione delle
risposte omeostatiche legate al ciclo giorno/notte.
La componente visiva più studiata è la via visiva primaria che va dalla retina al nucleo
genicolato laterale del talamo e da questo alla corteccia visiva primaria. Nell’ambito di questa
via diverse classi di neuroni codificano vari aspetti dell’informazione visiva: luminanza,
differenze spettrali, orientamento e movimento, che costituiscono ciò che noi vediamo.
L’elaborazione continua nelle vie corticali che si estendono oltre la corteccia visiva primaria e
raggiungono numerose aree visive nei lobi: occipitale, parietale e temporale.
PROIEZIONI CENTRALI DELLE CELLULE GANGLIARI RETINICHE
1. Via retino-genicolo-striata
2. Via retino-ipotalamica
3. Via retino-pretettale
4. Via retino-collicolare
RIFLESSO PUPILLARE ALLA LUCE: VIA RETINO-PRETETTALE
Questa via comprende proiezioni bilaterali dalla retina al pretetto e proiezioni bilaterali dal
pretetto al nucleo di Edinger-Westphal. I neuroni nel nucleo Edinger-Westphal terminano
ipsilateralmente nel ganglio ciliare e i neuroni nel ganglio ciliare innervano i muscoli costrittori
della pupilla. Si noti che gli assoni afferenti attivano entrambi i nuclei di Edinger-Westphal
attraverso i neuroni nel pretetto.
VIA VISIVA PRIMARIA: RETINO-GENICOLO-STRIATA
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Dopo essere entrate nella cavità cranica attraverso il foro ottico, le fibre del nervo ottico si
incrociano parzialmente nel chiasma ottico. L'incrociamento interessa solo le fibre che
provengono dalla metà nasale delle due retine. I tratti ottici, che dal chiasma ottico si portano
ai corpi genicolati laterali, risultano pertanto costituiti dalle fibre che provengono dalla metà
temporale della retina omolaterale e dalla metà nasale controlaterale. Nel tratto ottico di un
lato si trovano le fibre nervose che raccolgono le informazioni visive provenienti dalla metà del
campo visivo controlaterale. Questa disposizione è mantenuta anche nelle radiazioni ottiche
che si portano alla corteccia visiva primaria.
LA RAPPRESENTAZIONE RETINOTOPICA DEL CAMPO VISIVO
Proiezione di un'immagine sulla superficie della retina. Il passaggio dei raggi luminosi
attraverso la pupilla dell'occhio produce immagini che sulla retina sono capovolte e invertite
destra/sinistra.
Quadranti retinici e loro relazione con l'organizzazione di campi visivi monoculari e binoculari,
visti dalla superficie posteriore degli occhi. Le linee verticali e orizzontali tracciate attraverso il
centro della fovea definiscono i quadranti di ciascuna retina (in basso).
Linee analoghe tracciate attraverso il punto di fissazione definiscono i quadranti del campo
visivo (al centro). I diversi colori indicano le corrispondenze tra
i quadranti della retina e i quadranti del campo visivo.
CAMPO VISIVO DELL'OCCHIO
La macchia cieca è lo scotoma fisiologico, che è dovuto all'assenza di recettori per la
fuoriuscita delle fibre del nervo ottico.
La misura del campo visivo (statiche o cinetiche) prende il nome di perimetria o campimetria e
viene fatta per evidenziare perdite di sensibilità localizzate (scotomi), estese o riduzioni di
sensibilità.
Massima acuità visiva, nella macula lutea in corrispondenza della fovea e diminuisce verso la
periferia.
VISIONE BINOCULARE
I punti nella parte binoculare del campo visivo sinistro (B) cadono sulla retina nasale
dell'occhio sinistro e sulla retina temporale dell'occhio destro.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
I punti nella parte A differenza del nervo
binoculare del campo ottico, il tratto ottico
visivo destro (C) cadono contiene fibre
gangliari che
sulla retina nasale
originano da entrambi
dell'occhio destro e sulla gli occhi e
retina temporale rappresentano il
dell'occhio sinistro. campo visivo
I punti che si trovano nelle controlaterale.
porzioni monoculari dei Gli assoni delle cellule
campi visivo sinistro (A) e gangliari nella retina
nasale si incrociano
destro (D) cadono
nel chiasma ottico,
rispettivamente sulla mentre quelli della
retina nasale sinistra e retina temporale non
destra.Di conseguenza, le lo fanno. Il confine tra
informazioni dal campo cellule gangliari che
visivo sinistro vengono sporgono in senso
trasportate nel tratto ottico controlaterale o
ipsilaterale passa al
destro e le informazioni
centro della fovea e
dal campo visivo destro stabilisce il confine tra
vengono trasportate nel la retina nasale e
tratto ottico sinistro. temporale.
Una volta arrivate nelle aree corticali bersaglio, le terminazioni degli assoni che decorrono nel
tratto ottico, si dispongono ordinatamente a creare mappe accurate dell’emicampo
controlaterale. Nella via visiva primaria, la mappa dell’emicampo controlaterale ricostruita nel
nucleo genicolato laterale è conservata nelle proiezioni di questo nucleo alla corteccia striata.
La corteccia visiva primaria occupa gran parte del lobo occipitale. L'area della visione centrale
(la fovea) è rappresentata su una parte sproporzionatamente grande della porzione caudale
del lobo, mentre la visione periferica è rappresentata più anteriormente. Il campo visivo
superiore è rappresentato sotto la scissura calcarina, il campo inferiore sopra la scissura
calcarina.
DEFICIT DEL CAMPO VISIVO
Anopsia: ampio deficit del campo visivo
Scotoma: piccolo deficit del campo visivo
Esempi di deficit:
· Perdita della vista nell'occhio destro/sinistro
· Emianopsia bitemporale (eteronima): perdita della vista in entrambi gli occhi
delle metà esterne
· Emianopsia omonima sinistra/destra: perdita della vista in entrambi gli occhi
della metà sinistra/destra
· Quadrantanopsia superiore/inferiore sinistra/destra: perdita della vista in
entrambi gli occhi del quadrante in alto/basso a sinistra/destra
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Emianopsia omonima sinistra/destra con risparmio maculare: perdita della vista
in entrambi gli occhi della metà sinistra/destra senza la macula lutea (parte
centrale)
PERCORSO DELLA RADIAZIONE OTTICA FINO ALLA CORTECCIA STRIATA
Gli assoni che trasportano informazioni sulla porzione superiore del campo visivo si spostano
attorno al corno laterale del ventricolo nel lobo temporale (anello di Meyer) prima di raggiungere
il lobo occipitale.
Gli assoni che trasportano informazioni sulla parte inferiore del campo visivo viaggiano nel lobo
parietale.
I NEURONI NELLA CORTECCIA VISIVA PRIMARIA RISPONDONO SELETTIVAMENTE
ALL’ORIENTAMENTO DELLO STIMOLO
Esperimenti di David Hubel e Torsten Wiesel
La specificità delle proprietà del campo ricettivo dei neuroni nella corteccia striata (e oltre)
svolge un ruolo importante nel determinare gli attributi di base delle scene visive.
Un animale anestetizzato è dotato di lenti a contatto per focalizzare gli occhi su uno schermo,
dove è possibile proiettare immagini; un elettrodo extracellulare registra le risposte dei neuroni
corticali. I neuroni nella corteccia visiva primaria rispondono vigorosamente a una barra
luminosa orientata secondo un particolare angolo, mentre mostrano risposte deboli o nulle in
presenza di barre con altri orientamenti.
ORGANIZZAZIONE DELLA CORTECCIA VISIVA PRIMARIA
1. Aspetto della corteccia visiva primaria sottoposta a una colorazione istologica per mettere
in evidenza i corpi cellulari dei neuroni. Nei primati, lo strato 4 può essere ulteriormente
suddiviso (strati 4A, 4B, 4C).
2. Le cellule piramidali, con i loro prominenti dendriti apicali e basali sono il tipo cellulare più
numeroso nella neocorteccia; questi sono presenti in tutti gli strati ad eccezione del 4C. Lo
strato 4C è dominato da neuroni spinosi, i cui dendriti restano all’interno di questo strato.
3. Organizzazione laminare delle afferenze dal nucleo genicolato laterale (LGN). Gli assoni
dei neuroni residenti nel LGN terminano per la maggior parte negli strati 4C e 4A, mentre
innervano in maniera più dispersa gli strati 1, 2/3 e 6; nello strato 2/3 queste terminazioni sono
organizzate in «chiazze».
4. Organizzazione laminare delle principali connessioni intracorticali. I neuroni dello strato 4C
danno origine ad assoni che terminano in strati superficiali (4B e 2/3). Gli assoni di neuroni degli
strati 2/3 terminano abbondantemente nello strato 5. Gli assoni di neuroni dello strato 6
terminano nello strato 4C.
5. Organizzazione laminare di neuroni che proiettano a differenti bersagli. Le connessioni con
la corteccia extrastriata derivano principalmente dai neuroni degli strati 2/3 e 4B. Le proiezioni
discendenti al NGL derivano dai neuroni dello strato 6, mentre i neuroni che proiettano al
collicolo superiore risiedono nello strato 5.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
DOMINANZA OCULARE
L’integrazione, a livello della corteccia striata, dei segnali provenienti dai due occhi è alla base
della stereopsia. La stereopsia è la sensazione di profondità che ci deriva dal guardare oggetti
vicini con due occhi invece che con uno soltanto.
Far cells: si attivano in risposta alle disparità retiniche rilevate oltre il piano di fissazione.
Near cells: rispondono alle disparità retiniche relative a punti davanti al piano di fissazione.
Tuned zero: rispondono selettivamente a punti che giacciono sul punto di fissazione.
La conservazione delle risposte binoculari dei neuroni corticali dipende dalla normale attività dei
due occhi durante la prima vita postnatale. Tutto ciò che crea uno squilibrio nell'attività dei due
occhi, ad esempio l'allineamento anormale degli occhi durante l'infanzia (strabismo), può ridurre
in modo permanente l'efficacia di un occhio di attivare efficacemente i neuroni corticali e quindi
compromettere la capacità di utilizzare le informazioni binoculari come indizio di profondità. La
diagnosi precoce e la correzione dei problemi visivi è quindi essenziale per la normale funzione
visiva in età adulta.
NUCLEO GENICOLATO LATERALE
Nucleo genicolato laterale: strato magnocellulare (M), parvicellulare (P) e coniocellulare (K)
Le cellule M rispondono in modo transitorio alla presentazione di stimoli visivi, mentre le cellule
P rispondono in modo prolungato.
Le cellule gangliari P possono trasmettere informazioni sul colore, le cellule M no, perché le
aree periferiche e centrali dei loro campi recettivi sono attivate da differenti classi di coni.
Alcune cellule gangliari P ricevono al centro del loro campo recettivo input da coni sensibili di
lunghezza d'onda lunga ("rosso") mentre alla periferia del loro campo recettivo ricevono input
da coni di lunghezza d'onda media ("verde") e viceversa. Di conseguenza, le cellule P sono
sensibili alle differenze nelle lunghezze d'onda della luce che colpiscono il centro e la periferia
del loro campo recettivo.
Le cellule M non sono in grado di differenziare il tipo di segnale proveniente dai coni.
Le informazioni visive trasmesse dal flusso parvocellulare sono particolarmente importanti per
la visione ad alta risoluzione spaziale: l'analisi dettagliata della forma, delle dimensioni e del
colore degli oggetti.
Il sistema magnocellulare appare fondamentale per le attività che richiedono un'alta risoluzione
temporale, come la valutazione della posizione, della velocità e della direzione di un oggetto in
rapido movimento.
Il danno agli strati magnocellulari ha scarso effetto sull'acuità visiva o sulla visione dei colori, ma
riduce nettamente la capacità di percepire stimoli in rapido cambiamento. Al contrario, il danno
agli strati parvocellulari non ha alcun effetto sulla percezione del movimento ma compromette
gravemente l'acuità visiva e la percezione del colore.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
L’UDITO
L’apparato uditivo trasduce vibrazioni meccaniche trasformandole in sensazioni sonore.
suono = è una vibrazione che insorge da uno strumento e poi si propaga per mezzo dell’aria
FISICA DEL SUONO
In termini fisici, il suono si riferisce alle onde di pressione generate dalle molecole d'aria che
vibrano (erroneamente, il termine suono viene usato più comunemente per riferirsi a una
percezione uditiva). Le onde sonore sono molto simili alle increspature che si irradiano verso
l'esterno quando una pietra viene lanciata in una pozza d'acqua. Tuttavia, invece di verificarsi
su una superficie bidimensionale, le onde sonore si propagano in tre dimensioni, creando
alternate cicliche compressioni e rarefazione.
Importante è il mezzo, senza il quale il suono non esisterebbe.
Come tutti i fenomeni ondulatori, le onde sonore hanno delle caratteristiche principali: forma
d'onda, ampiezza (solitamente espressa come decibel, dB abbreviato) e frequenza (espressa
in cicli al secondo o Hertz, abbreviata Hz).
Per l’ orecchio umano, l'ampiezza e la frequenza di una variazione della pressione sonora
corrispondono rispettivamente al volume e all'intonazione.
L’oscillazione dei rebbi del diapason si trasmette alle molecole d’aria circostanti inducendo
una serie di cicliche compressioni e rarefazioni.
Questi fenomeni possono essere descritti in un grafico che riporta l’andamento della pressione
nel tempo e nello spazio e permette di quantificare alcuni parametri:
· Ampiezza: (dalla massima rarefazione alla massima compressione) corrisponde al
contenuto d’energia dello stimolo sonoro e quindi alla sua intensità sensoriale (dB);
· Frequenza: numero di oscillazioni nell’unità di tempo (altezza o tonalità dello stimolo
sonoro; a bassa frequenza sono percepiti come gravi ad alta frequenza come acuti);
· Forma d’onda: permette di classificare diversi tipi di stimoli sonori;
La vibrazione dei rebbi di un diapason induce una serie di compressioni e rarefazioni
cicliche delle molecole dell'aria circostante, che possono essere descritte da un grafico nel
quale sono riportate le variazioni pressorie nel tempo o nello spazio.
I suoni prodotti dagli strumenti musicali o dalla voce umana sono costituiti da più frequenze
che si ripetono in maniera ciclica costante. Infine, i rumori sono costituiti da diverse
frequenze che si susseguono senza alcun ordine o andamento periodico.
I toni puri, prodotti dai diapason o dagli audiometri, sono oscillazioni sinusoidali,
caratterizzate da un'unica frequenza.
SPETTRO DELLE FREQUENZE UDIBILI DALL’UOMO
L’uomo può udire frequenze nel range 20Hz-20kHz. Si noti come, in tutti i casi, l'intensità
dello stimolo (in decibel) necessaria per evocare una certa intensità di sensazione sia
minore per le frequenze intermedie (all'incirca fra 1.000 e 4.000 Hz) e aumenti spostandosi
verso le alte o le basse frequenze.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
ORECCHIO ESTERNO
L'orecchio esterno è composto da: pinna, conca (o padiglione auricolare) e meato uditivo,
raccoglie l’energia sonora e la convoglia verso il timpano o membrana timpanica.
Rende gli esseri umani particolarmente sensibili a frequenze comprese tra 2 e 5 kHz, e
spiega perché le perdite di udito siano intorno a questo ambito di frequenza in seguito a
esposizione a forti rumori. La sensibilità del sistema uditivo umano in questa gamma di
frequenze sembra sia correlata alla percezione del linguaggio.
Altra importante funzione della pinna e padiglione auricolare è quella di filtrare le diverse
frequenze allo scopo di fornire informazioni relative all’altezza in cui si trova la sorgente
sonora. La pinna ha una conformazione per trasmettere un maggior numero di componenti
ad alta frequenza da una fonte sonora elevata che dalla stessa fonte è situata a livello
dell’orecchio.
ORECCHIO MEDIO
è costituito da:
· Membrana timpanica;
· Catena dei tre ossicini: martello, incudine,
staffa;
· Muscoli tensore del timpano e stapedio;
(aria orecchio esterno; acqua orecchio interno)
La funzione principale è quella di adattare
l’impedenza relativamente bassa dei suoni trasmessi per via aerea alla più elevata
impedenza del liquido dell’orecchio interno.
PRINCIPALE FUNZIONE DELL’ORECCHIO MEDIO
Due processi meccanici e un processo nervoso avvengono all'interno dell'orecchio per
realizzare questo grande aumento di pressione.
Il primo aumento di pressione avviene convogliando la forza che agisce sul timpano del
diametro relativamente ampio sulla finestra ovale il cui diametro è molto ridotto, dove le
ossa dell’orecchio medio entrano in contatto con l’orecchio interno.
Il secondo vantaggio meccanico è realizzato dal sistema di leva costituito dalla catena degli
ossicini, che aumentano l’ampiezza della vibrazione e che collegano il timpano alla finestra
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
ovale dove la platina della staffa si inserisce come un pistone e trasmette le vibrazioni ai
liquidi dell’orecchio interno.
Questo meccanismo è dotato di un sensibile sistema di regolazione, costituito da due
piccoli muscoli: il tensore del timpano e lo stapedio, innervati dal V e dal VII,
rispettivamente. Il primo si inserisce sulla testa del martello e aumenta la tensione del
timpano, lo stapedio si inserisce sul collo della staffa e contraendosi esercita una trazione
sulla staffa riducendone l’affondamento nella finestra ovale.
Nell’insieme i due muscoli aumentano la rigidità della catena timpano ossiculare riducendo
la conduzione degli stimoli sonori. Protezione dell’orecchio da stimoli troppo intensi.
ORECCHIO INTERNO: LA COCLEA
La coclea dell'orecchio interno è la struttura più affascinante e importante delle vie
acustiche, poiché è nella coclea che l'energia delle onde di pressione generate dai suoni
viene trasformata in impulsi nervosi.
La coclea non solo amplifica le onde sonore e le converte in impulsi nervosi, ma funge
anche da analizzatore meccanico di frequenza, decomponendo le forme d'onda acustiche
complesse in componenti più semplici.
Molte caratteristiche della percezione uditiva derivano direttamente dalle proprietà fisiche
della coclea. La coclea (dal latino «chiocciola") è una minuscola struttura a spirale (circa 10
mm di larghezza) che, se venisse despiralizzata, formerebbe un canale lungo circa 35 mm.
Sia la finestra ovale che la finestra rotonda, un’altra regione che circonda la coclea (regioni
in cui l'osso è assente), si trovano all'estremità basale di questo canale.
> cosa succede quando arriva un’onda sonora:
Quando arriva un'onda sonora, Il compito del padiglione auricolare è quello di fare da
antenna = Captare l'onda sonora. Una volta capitata, viene incanalata all'interno del meato
uditivo il quale raccoglie l'energia sonora e la convoglia verso il timpano potevi, questa
vibrazione fa vibrare il timpano e Fa muovere a loro volta i tre ossicini: la staffa (va
immaginata come un pistone) preme su una parte della coclea, la quale si chiama finestra
ovale. Quando la staffa preme, come una sorta di pistone, sulla finestra ovale della coclea
la pressione genera un movimento, fa muovere il fluido contenuto all’interno della coclea.
All’interno della coclea ci stanno i vari recettori uditivi che sono delle cellule che si
chiamano cellule cigliate.
STRUTTURA DELL'ORECCHIO INTERNO
La coclea è un canalicolo spiraliforme. Al suo interno, le strutture del labirinto membranoso
delimitano tre scale che corrono parallele per tutta la lunghezza. La disposizione relativa
delle tre scale: la scala vestibolare, la scala timpanica e la scala media. Quest'ultima, di
sezione triangolare, è delimitata da tre membrane: la membrana basilare, sulla quale si
trova l'organo di Corti, la separa dalla scala timpanica; la membrana di Reissner la separa
dalla scala vestibolare; la stria vascolare la delimita verso la faccia esterna della coclea.
La scala vestibolare parte dalla finestra ovale nella quale è inserita la staffa, sale fino
all’apice del canalicolo cocleare dove, attraverso l’elicotrema, comunica con la scala
timpanica. Questa termina alla base della coclea in corrispondenza della finestra rotonda,
un forellino nell’osso temporale chiuso da una membrana elastica. Fra queste due scale si
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
trova la scala media. Le scale timpanica e vestibolare contengono un fluido detto perilinfa
che si mescola nel elicotrema, la scala media contiene endolinfa.
L’apparato recettoriale vero e proprio è costituito dall’organo di Corti, un ispessimento
dell’epitelio che copre la membrana basilare, il quale contiene oltre a cellule di sostegno i
recettori acustici: le cellule ciliate. Una conseguenza di questa disposizione strutturale è
che il movimento verso l'interno della finestra ovale sposta il fluido dell'orecchio interno,
facendo sporgere leggermente la finestra rotonda e deformando la partizione cocleare.
ORGANO DI CORTI
L'organo di Corti è costituito da un ispessimento dell'epitelio che ricopre la membrana
basilare. L’organo di Corti è il posto dove sono collocate le cellule cigliate. Queste cellule
sono disposte in tre file esterne e una sola interna. Le cellule cigliate esterne toccano con
la membrana superiore; è una membrana gelatinosa e si chiama membrana tettoria.
CELLULE CILIATE ESTERNE ED INTERNE
Le cellule ciliate interne sono i veri e propri recettori sensoriali. Le fibre delle cellule ciliate
esterne sono tutte costituite da assoni efferenti provenienti dal complesso olivare superiore.
Svolgono funzione di modulazione dei movimenti della membrana basilare e costituiscono
un'importante componente cocleare.
Il movimento della membrana basilare, crea un'onda che genera uno sfregare tra le cellule
cigliate esterne e la m. tettoria. Per via di questo movimento le cellule cigliate esterne
toccano la membrana tettoria, le loro ciglia si muovono con una direzione preferenziale che
porta a una depolarizzazione poi della cellula e invia il segnale nervoso al sistema nervoso.
TONOTOPIA
Georg von Békésy dimostrò che una membrana che varia sistematicamente in larghezza e
flessibilità vibra in maniera massimale nelle diverse posizioni in funzione della frequenza
dello stimolo.
Utilizzando modelli tubolari e coclee umane prelevate da cadaveri, egli scoprì che uno
stimolo acustico avvia un'onda viaggiante nella coclea della stessa frequenza nella coclea,
che si propaga dalla base verso l'apice della membrana basilare, crescendo in ampiezza e
rallentando la velocità fino a raggiungere il punto massimo di spostamento. Questo punto
massimo di spostamento è determinato dalla frequenza del suono e persiste vibrando allo
stesso modo per tutta la durata del suono. I punti che rispondono alle alte frequenze sono
alla base della membrana basilare dove è più rigida, i punti che rispondono alle basse
frequenze sono all'apice, dando origine a una rappresentazione topografica delle frequenze
(cioè alla tonotopia).
ONDE CHE VIAGGIANO LUNGO LA COCLEA
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Le misurazioni della vibrazione di diverse parti della membrana basilare, nonché le velocità
di scarica delle singole fibre nervose uditive che terminano lungo la sua lunghezza,
mostrano che entrambe queste caratteristiche sono altamente sintonizzate; cioè,
rispondono più intensamente a un suono di una frequenza specifica.
La sintonizzazione della frequenza all'interno dell'orecchio interno è in parte attribuibile alla
geometria della membrana basilare, che è più ampia e flessibile all'estremità apicale e più
stretta e rigida all'estremità basale. La caratteristica di un tale sistema è che,
indipendentemente da dove viene fornita energia, il movimento inizia sempre dalla parte
rigida (cioè la base) e poi si propaga alla parte più flessibile (cioè l'apice).
MOVIMENTO DELLA MEMBRANE BASILARE
Il movimento della membrana basilare determina il piegamento di sottili processi chiamati
stereociglia che sporgono all’estremità superiore delle cellule ciliate. Il punto di appoggio
della membrana basilare non è allineato al punto di appoggio della membrana tettoria, in
modo che quando la membrana basilare viene spostata, la membrana tettoria scivola
sull’estremità superiore delle cellule ciliate determinando un piegamento delle stereociglia.
Il movimento dell'onda viaggiante avvia la trasduzione sensoriale spostando le cellule
ciliate che si trovano in cima alla membrana basilare.
CELLULE CILIATE
La cellula ciliata è una conquista evolutiva che risolve il problema di trasformare l’energia
vibrazionale in un segnale elettrico. Sono in grado di rilevare movimenti delle dimensioni di
un atomo e di rispondere nel giro di pochi millisecondi. Sono capaci di adattarsi con rapidità
a stimoli costanti, permettendo all’ascoltatore di distinguere determinati segnali dai rumori
di fondo.
STRUTTURA E FUNZIONE DEL FASCIO DI CIGLIA NELLE CELLULE CILIATE DELLA
COCLEA
I fasci di ciglia qui raffigurati sono simili a quelli della coclea, ad eccezione della presenza
del chinociglio, che nella coclea dei mammiferi scompare poco dopo la nascita.
Chinociglio:
A. Progressivo aumento in lunghezza delle stereociglia in direzione del chinociglio.
B. Sezione trasversale del fascio di ciglia vestibolare che mostra in alto la
disposizione 9+2 dei microtubuli del chinociglio, rispetto alla struttura più
semplice delle stereociglia, costituite da fasci di filamenti di actina.
C. Fascio di cellule cigliate esterne della coclea. È da notare la lunghezza graduata
delle stereociglia e l’assenza del chinociglio.
D. Tip link che connettono stereociglia adiacenti. Si ritiene siano i legami meccanici
che causano l’apertura e la chiusura dei canali di trasduzione.
TIP LINK
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Nell’insieme , l’apparato funziona come una molla: se viene messo in trazione, la porta del
canale si apre e gli ioni possono fluire attraverso il poro; se viene rilasciato, la porta si
chiude e il canale non è più pervio.
Al contrario un movimento nella direzione opposta , avvicina le punte delle stereociglia,
riduce la tensione del filamento proteico e determina la chiusura del canale di trasduzione.
La disposizione a scalare delle cellule di diversa lunghezza fa sì che quando il ciuffo si
flette verso la fila delle stereociglia più lunghe la distanza fra le punte delle stereociglia
delle file successive aumenta, il tip link viene stirato e le porte dei canali si aprono.
Tip link: filamento proteico che connette lo stereociglio adiacente appartenente alla fila
successiva. Alle estremità è collegato con una proteina canale, detta canale di trasduzione,
situata nella membrana dello stereociglio .
TRASDUZIONE ELETTROMECCANICA MEDIATE DALLE CELLULE CILIATE
Quando il fascio di ciglia si piega verso lo stereociglio più alto, si aprono canali ionici
specifici per cationi in prossimità delle punte delle stereociglia, che consentono l’entrata di
K⁺.La conseguente depolarizzazione causa l’apertura dei canali per il Ca²⁺ voltaggio
dipendenti nel soma cellulare, permettendo al Ca²⁺ di entrare ed innescare l’esocitosi del
neurotrasmettitore nelle terminazioni del nervo acustico.
PROPRIETÀ FUNZIONALI DEI NEURONI DEL GANGLIO DI CORTI
La frequenza caratteristica dei neuroni del ganglio di Corti è determinata dalla loro
relazione esclusiva con una sola cellula recettrice (codice di posizione).
L'attività elettrica di questi neuroni è spesso in relazione con la fase dello stimolo (il
potenziale d'azione insorge più facilmente in corrispondenza del massimo dell'onda sonora,
quando il recettore è depolarizzato).
L'attività (frequenza dei potenziali d'azione) di un neurone del ganglio di Corti è
proporzionale all'intensità dello stimolo. In particolare, si osserva un brusco incremento in
coincidenza con l'inizio dello stimolo (burst); poi l'attività si assesta su un livello più basso
(plateau), che resta costante per tutta la durata della stimolazione.
Un singolo neurone può misurare variazioni di intensità in un ambito di circa 40 dB. L'intera
gamma delle intensità udibile (120 dB) è segnalata attraverso il reclutamento di più neuroni
dotati di diverse soglie di attivazione (codice di popolazione).
PROPRIETÀ DI RISPOSTA DELLE FIBRE DEL NERVO ACUSTICO
Curve tonali di frequenza registrate da sei diverse fibre del nervo acustico. Ogni grafico
rappresenta, per tutte le frequenze a cui la fibra risponde, il livello minimo di intensità
sonora necessario per aumentare la frequenza di scarica della fibra al di sopra del suo
livello di scarica spontanea. Il punto più basso del tracciato rappresenta l'intensità sonora
più debole alla quale il neurone risponde. La frequenza in questo punto è chiamata
frequenza caratteristica del neurone.
Curve tonali di frequenza delle fibre del nervo acustico sovrapposte e allineate con i loro
relativi punti di innervazione lungo la membrana basilare (rappresentata come una linea
nera all'interno della coclea srotolata).
INNERVAZIONE DELLE CELLULE CILIATE DELL’ORGANO DI CORTI
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
La grande maggioranza degli assoni sensoriali nella coclea trasporta segnali dalle cellule
ciliate interne, ognuna delle quali costituisce l'unico input per una media di 10 assoni.
Alcuni assoni sensoriali di piccolo calibro trasmettono informazioni dalle cellule ciliate
esterne. Gli assoni efferenti innervano in gran parte le cellule ciliate esterne e lo fanno
direttamente. Al contrario, l'innervazione efferente delle cellule ciliate interne è scarsa e si
verifica sui terminali degli assoni sensoriali.
LA VIA UDITIVA PRIMARIA
Il primo stato dell'elaborazione uditiva centrale nel cervello avviene nel nucleo cocleare
(parte rostrale del bulbo nel mesencefalo, formato da tre parti distinte), il bersaglio iniziale
degli assoni nel nervo uditivo che portano l’informazione generata nella membrana
basilare. È la sola stazione della via acustica che riceve informazioni monoaurali
dall’orecchio ipsilaterale.La proiezione che origina dai nuclei cocleari è complessa e
costituita da vie parallele distinte.
La via acustica comprende diversi nuclei
del tronco encefalico come:
· Corteccia uditiva primaria (lobo
temporale)
· Complesso genicolato del talamo
· Collicolo inferiore
· Nucleo del lemnisco laterale
· Complesso olivare superiore
Questa organizzazione evidenzia due punti
importanti:
1. il sistema uditivo comporta diversi
percorsi paralleli
2. le informazioni provenienti da ciascun
orecchio raggiungono entrambi i lati del
sistema, anche a livello del tronco
encefalico.
L’elevato grado di connessione bilaterale
implica che un danno alle strutture uditive
centrali non si manifesta mai con sordità
monoaurale. La perdita dell’udito da un solo
orecchio indica fortemente un danno
periferico monolaterale, all’orecchio medio o
interno oppure al nervo acustico.
All'interno del nucleo cocleare, ogni fibra
nervosa uditiva si dirama, inviando un ramo ascendente al nucleo cocleare anteroventrale
e un ramo discendente al nucleo cocleare postero ventrale e al nucleo cocleare dorsale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
L'organizzazione tonotopica della coclea viene mantenuta nelle tre parti del nucleo
cocleare, ognuna delle quali contiene diverse popolazioni di cellule con proprietà
abbastanza diverse.
LOCALIZZAZIONE DEI SUONI
La funzione meglio conosciuta e più intensamente studiata tra quelle mediate dai nuclei
uditivi del tronco encefalico, è la corretta localizzazione dei suoni.
Gli umani usano almeno due diverse strategie per localizzare la posizione orizzontale delle
sorgenti sonore, a seconda delle frequenze nello stimolo.
Per frequenze inferiori a 3 kHz (che possono essere seguite secondo la modalità di
sincronizzazione di fase), le differenze di tempo interaurali sono utilizzate per localizzare la
fonte sonora; al di sopra di 3 kHz vengono utilizzate differenze di intensità interaurale.
Percorsi paralleli che originano dal nucleo cocleare rispondono a ciascuna di queste
strategie di localizzazione dei suoni.
La capacità umana di rilevare differenze di tempo interaurali è notevole. Le più lunghe
differenze di tempo interaurale, che sono prodotte da suoni che sorgono direttamente
lateralmente a un orecchio, sono dell'ordine di soli 700 µs (valore dato dalla larghezza della
testa diviso per la velocità del suono nell'aria, circa 340 m/s ). Gli esseri umani possono
rilevare differenze di tempo interaurali di appena 10µs; due suoni presentati attraverso
auricolari, separati da differenze di tempo interaurali così piccole sono percepiti come
provenienti dalla direzione del lato dell'orecchio stimolato per primo. Questa sensibilità si
traduce in una precisione di localizzazione del suono di circa 1 grado.
RAPPRESENTAZIONE DELLO SPAZIO UDITIVO ESTERNO
La rappresentazione dello spazio uditivo esterno viene costruita lungo la via acustica
centrale analizzando le differenze di tempo e intensità con cui ogni stimolo arriva alle due
orecchie.
Uno stimolo proveniente da una sorgente sonora posta a sinistra del soggetto giungerà
prima all'orecchio sinistro rispetto a quello destro. Inoltre, dato che la testa riflette una parte
dello stimolo, esso giungerà anche leggermente attenuato all'orecchio destro. Questo
effetto di ombra sonora è maggiore per le alte frequenze e, perciò, le differenze di intensità
binaurale sono il parametro fondamentale per la localizzazione spaziale di stimoli ad alta
frequenza.
TEMPO E DIREZIONE DELLO STIMOLO
Nell'oliva superiore, vengono confrontate le minuscole differenze di tempi e intensità del
suono in ciascun orecchio, per determinare la direzione da cui proviene il suono.
Il nervo acustico trasporta il segnale nel tronco cerebrale e si ramifica nel nucleo cocleare.
Le cellule del nucleo cocleare ventrale proiettano quindi verso un gruppo di nuclei nel
midollo chiamato oliva superiore.
L'oliva superiore porta poi verso il collicolo inferiore attraverso un tratto di fibre chiamato
lemnisco laterale.
POSIZIONE DELLA FONTE SONORA SULLA BASE DELLA DIVERSA INTENSITÀ
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Frequenze superiori ai 2kHz, la testa umana inizia ad agire come un ostacolo acustico,
perché le lunghezze d’onda dei suoni sono troppo brevi per piegarsi attorno a essa (ombra
acustica). Queste differenze di intensità forniscono un secondo indizio sulla posizione di un
suono. I circuiti che provvedono a questo sono l’oliva superiore laterale (LSO) e il nucleo
mediale del corpo trapezoide (MNTB).
I neuroni LSO ricevono l'eccitazione diretta dal nucleo cocleare omolaterale; L'input dal
nucleo cocleare controlaterale viene trasmesso attraverso interneuroni inibitori nell'MNTB.
1. Lo stimolo che giunge all’orecchio sinistro con maggiore intensità attiva la LSO
di sinistra.
2. Questo stimolo inibisce la LSO di destra tramite un interneurone del MNTB.
3. L’attivazione sinistra è maggiore dell’inibizione destra, con il risultato di
un’attivazione netta verso i centri superiori.
4. L’attivazione sinistra è maggiore dell’inibizione destra, con il risultato di
un’attivazione netta a destra e assenza di segnali verso i centri superiori.
QUALITÀ DEL SUONO
Il secondo flusso di informazioni inizia nel nucleo cocleare dorsale.
Il nervo acustico trasporta il segnale nel tronco cerebrale e si ramifica nel nucleo cocleare.
Questa via proietta direttamente al collicolo inferiore e anche attraverso il lemnisco laterale.
Il nucleo cocleare dorsale, con circuiti piuttosto complessi, analizza le minuscole differenze
di frequenza che rendono il suono diverso esempio fonemi.
PERCORSI MONOAURALI DAL NUCLEO COCLEARE AL LEMNISCO LATERALE: INIZIO
E DURATA
Un secondo importante gruppo di vie che partono dal nucleo cocleare oltrepassa l'oliva
superiore e termina nei nuclei del lemnisco laterale sul lato controlaterale del tronco
encefalico. Queste particolari vie trasportano informazioni acustiche che arriva ad un solo
orecchio e sono quindi definite monoaurali.
Alcune cellule dei nuclei del lemnisco laterale segnalano l'inizio del suono,
indipendentemente dalla sua intensità o frequenza.
Altre cellule dei nuclei del lemnisco laterale elaborano altri aspetti temporali del suono,
come la durata.
Il ruolo preciso svolto da queste vie nell'elaborazione delle caratteristiche temporali del
suono non è ancora noto. Come le vie dei nuclei olivari superiori, le vie che hanno origine
dai nuclei del lemnisco laterale convergono a livello del mesencefalo.
I PROCESSI DI INTEGRAZIONE A LIVELLO DEL COLLICOLO INFERIORE
Le vie acustiche che salgono passando attraverso il complesso dei nuclei olivari e
lemniscali, insieme ad altre proiezioni che nascono direttamente dal nucleo cocleare,
proiettano al collicolo inferiore il centro acustico del mesencefalo.
Nell'esaminare come avviene l'integrazione nel collicolo inferiore, è di nuovo utile ricorrere
al meccanismo uditivo più completamente analizzato, il sistema binaurale per localizzare il
suono. Come già notato, non esiste una mappa topografica dello spazio sulla superficie dei
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
recettori acustici; di conseguenza, la percezione dello spazio uditivo deve in qualche modo
essere sintetizzata dai circuiti delle aree inferiori del tronco encefalico e del mesencefalo.
Esperimenti nel barbagianni, un animale straordinariamente abile nel localizzare i suoni,
mostrano che la convergenza degli input binaurali nel mesencefalo produce qualcosa di
completamente nuovo rispetto alla periferia, vale a dire una rappresentazione topografica
calcolata dello spazio uditivo.
I neuroni all'interno di questa mappa spaziale uditiva nel collicolo rispondono meglio ai
suoni originati in una specifica regione di spazio e quindi riescono a stabilire una
localizzazione preferenziale sia in verticale che in orizzontale, o azimut. Sebbene nell’uomo
non siano ancora state trovate mappe comparabili dello spazio uditivo, gli umani hanno una
chiara percezione delle componenti sia di altezza sia azimutale della localizzazione di un
suono, suggerendo di possedere una rappresentazione nervosa dello spazio uditivo.
VIA COLLICOLO-COMPLESSO GENICOLATO
Dal collicolo inferiore, i flussi di informazioni uditive che arrivano dall’oliva superiore e dalle
altre vie parallele arrivano al talamo sensoriale.
Il nucleo uditivo del talamo è il complesso genicolato mediale.
Il complesso genicolato mediale proietta alla corteccia uditiva primaria, situata nel lobo
temporale.
IL TALAMO ACUSTICO
Il complesso genicolato mediale (MGC) del talamo è una stazione obbligatoria di transito e
di trasmissione di tutte le informazioni uditive ascendenti destinate alla corteccia.
La maggior parte dell'input all'MGC deriva dal collicolo inferiore, sebbene alcune fibre
acustiche provenienti dalle aree inferiori dal tronco encefalico aggirino il collicolo inferiore e
raggiungano direttamente il talamo acustico.
L'MGC è distinto in varie parti, tra cui la parte ventrale, che funziona come principale
stazione di ricezione e trasmissione delle informazioni a livello talamo-corticale e le parti
dorsale e mediale, che sono disposte come una fascia attorno alla divisione ventrale.
In alcuni mammiferi, l’organizzazione rigorosamente tonotopica delle aree inferiori del
tronco encefalico viene realizzata mediante convergenza sui neuroni dell’ MGC,
generando:
· risposte specifiche a determinate combinazioni di frequenza;
· risposte specifiche non solo alle combinazioni di frequenza ma anche a specifici
intervalli di tempo tra le due frequenze.
In sintesi, i neuroni nel complesso genicolato mediale ricevono afferenze convergenti da
percorsi spettrali e temporali separati. In virtù delle afferenze convergenti che riceve,
questo complesso media l’integrazione di specifiche combinazioni spettrali e temporali di
suoni.
Si noti che l'elaborazione del parlato richiede certamente sensibilità sia della combinazione
spettrale che temporale.
LA CORTECCIA UDITIVA UMANA
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
L'obiettivo finale delle informazioni uditive afferenti è la corteccia uditiva. Sebbene la
corteccia uditiva abbia una serie di suddivisioni, si può fare una grande distinzione tra
un'area primaria (A1) e aree periferiche, o cintura (BELT).
La corteccia uditiva primaria (A1) si trova sul giro temporale superiore nel lobo temporale e
riceve afferenze punto-punto dalla parte ventrale del complesso genicolato mediale; quindi
contiene una precisa mappa tonotopica.
Le aree periferiche della corteccia uditiva ricevono afferenze più diffuse dalle aree
circostanti il complesso genicolato mediale, oltre che da A1, e quindi presentano una
organizzazione tonotopica meno precisa.
Poiché le frequenze sono organizzate tonotopicamente per tutta la lunghezza della
membrana basilare si assume che A1 comprenda una mappa tonotopica, così come la
maggior parte delle strutture uditive ascendenti tra la coclea e la corteccia.
ORGANIZZAZIONE FUNZIONALE
Perpendicolarmente all'asse delle frequenze della mappa tonotopica ci sono strie di
neuroni che vengono eccitati da entrambe le orecchie e per questa ragione vengono
chiamate cellule EE, inframmezzate da aree occupate da neuroni che vengono eccitati da
un orecchio e inibiti dall'altro orecchio cellule EI.
Le strie EE ed EI si alternano (una disposizione che ricorda le colonne di dominanza
oculare in V1).
La corteccia uditiva ovviamente fa molto di più che fornire una mappa tonotopica e
rispondere in modo diverso alla stimolazione ipsi- e controlaterale. Sebbene i tipi di
elaborazione sensoriale che si verificano nella corteccia uditiva non siano ben compresi, è
probabile che siano importanti per l'elaborazione dei suoni naturali di ordine superiore, in
particolare quelli utilizzati per la comunicazione.
I neuroni nella corteccia uditiva sono "sensibili alla combinazione" in quanto rispondono a
specifiche combinazioni temporali e spettrali di suoni naturali rispetto al rumore. Ciò
suggerisce che l'elaborazione corticale è in parte dedicata alla rilevazione di particolari
vocalizzazioni intraspecifiche.
RAPPRESENTAZIONE TONOTOPICA E ORGANIZZAZIONE COLONNARE NELLA
CORTECCIA UDITIVA PRIMARIA
Nell'area A1 dei mammiferi, le frequenze sono rappresentate in maniera progressiva lungo
una serie di moduli paralleli che si susseguono lungo la superficie corticale. Nella stessa
area è presente un altro sistema di moduli alternati, definiti dalle caratteristiche della
risposta dei neuroni alle informazioni provenienti dalle due orecchie. Nei moduli di
sommazione (EE), i neuroni sono eccitati da entrambe le afferenze, nei moduli di
soppressione (EI) sono eccitati dalle proiezioni provenienti dall'orecchio controlaterale e
inibiti da quelle ipsilaterali. Il sistema dei moduli di frequenza e quello dei moduli di
sommazione/soppressione sono disposti ortogonalmente, in modo da definire colonne
corticali nelle quali i neuroni hanno in comune la frequenza caratteristica e le modalità di
risposta alle informazioni binaurali.
APPARENZA ACUSTICA E VESTIBOLARE
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Nella colonna delle afferenze somatiche speciali troveremo sia i nuclei cocleari e sia i
nuclei vestibolari. Tutte e due fanno riferimento al nucleo vestibolare cocleare (8° nervo
cranico).
-nuclei cocleari → riportano le afferenze di tipo uditivo
-nuclei vestibolari → inerenti al senso dell’equilibrio
NUCLEI COCLEARI
Li possiamo trovare in una sezione trasversale a livello di Ponte; troveremo:
-Nucleo cocleare ventrale | Riceveranno le afferenze provenienti
-Nucleo copiare dorsale | dal ganglio di Corti
Tutte le afferenze provenienti da questo ganglio faranno sinapsi a livello di nucleo cocleare
ventrale, in alcuni casi queste afferenze cederanno una collaterale anche per il nucleo
cocleare dorsale.
da questi nuclei partiranno diverse fibre:
- per quanto riguarda il n.c.v. una piccola componente delle fibre che si porteranno
ventralmente andrà a fare sinapsi nel nucleo olivare omolaterale punto e virgola mentre la
maggior parte andranno a fare sinapsi nel nucleo olivare Superiore controlaterale.
> queste fibre che si porteranno ventralmente formeranno quella che verrà chiamata stria
acustica ventrale
→ Vi sarà sempre che si originerà nel nucleo cocleare ventrale interiore stria = stria
acustica intermedia
> le fibre che originano dal nucleo si porteranno In un primo momento dorsalmente quindi
the correndo dietro a quello che è il peduncolo cerebellare Inferiore e passando Attraverso
al nucleo cocleare dorsale
→ A livello più o meno del 4 ventricolo si porteranno nuovamente mentalmente per fare sti
ragazzi nel nucleo olivare Superiore controlaterale.
-Per quanto riguarda il n.c.d vi saranno anche qui delle fibre che formeranno la stria
acustica dorsale.
funzioni:
-nucleo cocleare ventrale: darà informazioni inerenti all’intensità
-nucleo cocleare dorsale: darà informazioni inerenti alla frequenza
-complesso olivare superiore: darà informazioni inerenti alla posizione
ORGANIZZAZIONE FUNZIONALE
I neuroni sensibili alla combinazione sono sintonizzati su più di una frequenza e sono
specializzati per riconoscere suoni complessi specie specifici e per estrarre informazioni
che sono fondamentali per la sopravvivenza.
Questa sensibilità alle combinazioni di semplici elementi sonori sembra essere una
proprietà universale dei neuroni per la percezione di suoni complessi da parte di molte
specie animali, come rane, pipistrelli ,uccelli, primati non umani e umani.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Quei suoni particolarmente importanti per la comunicazione intraspecifica spesso hanno
una struttura temporale altamente ordinata. Nell'uomo, il miglior esempio di segnali che
variano nel tempo è la parola, in cui diverse sequenze fonetiche sono percepite come
sillabe e parole distinte.
L'area di Wernicke, che è fondamentale per la comprensione del linguaggio umano, si trova
all'interno dell'area uditiva secondaria.
Gli studi su pazienti umani con danno bilaterale alla corteccia uditiva rivelano anche gravi
problemi nell'elaborazione dell'ordine temporale dei suoni.
Sembra quindi probabile che specifiche diverse regioni della corteccia uditiva umana siano
specializzate:
· Per l'elaborazione di suoni vocali elementari
· Segnali acustici temporalmente complessi, come la musica
ORGANIZZAZIONE E PRINCIPALI CONNESSIONI DELLE AREE CORTICALI UDITIVE
L'elaborazione corticale delle informazioni uditive è effettuata da diverse aree organizzate
in maniera gerarchica e parallela.
Le aree centrali (core area), che ricevono le afferenze talamiche (MGd, MGv),
comprendono la corteccia uditiva primaria (A1) e due aree adiacenti, definite rostrale (R) e
caudo mediale (CM).
Accanto a queste si trovano tre aree periferiche (belt area), denominate area anterolaterale
(AL), mediolaterale (ML) e caudolaterale (CL).
Di qui il flusso dell'informazione uditiva segue due principali vie parallele:
· La via dorsale, diretta al lobo frontale (PFC) attraverso la corteccia parietale
posteriore (PP) è maggiormente coinvolta nella localizzazione spaziale degli stimoli
sonori ("Dove").
· La via ventrale giunge ad altre aree della corteccia prefrontale (PFC) procedendo
attraverso le regioni anteriori della corteccia temporale (parabelt area, PB; aree
temporali T2/T3). Questa via sarebbe responsabile del riconoscimento della natura
dello stimolo uditivo ("Cosa").
I SENSI CHIMICI
I sensi chimici sono il sistema olfattivo, gustativo e chemiosensoriale del trigemino, che
hanno in comune la capacità di riconoscere gli stimoli chimici presenti nell’ambiente.
Tutti e tre questi sistemi chemiosensoriali dipendono da recettori nella cavità nasale, nella
bocca o sul viso che interagiscono con le rispettive molecole presenti nell’ambiente,
generando potenziali di azione e trasmettendo così informazioni sugli stimoli chimici ad
appropriate regioni del sistema nervoso centrale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
OLFATTO
Il sistema olfattivo rileva molecole disperse nell'aria chiamate sostanze odorose.
Nell'uomo, gli odori forniscono informazioni su sostanze chimiche presenti nel proprio corpo
e nelle altre persone, negli animali, nelle piante e aiuta inoltre a identificare i cibi e le
sostanze nocive presenti nell’ambiente. Quindi le informazioni olfattive possono influenzare
il comportamento alimentare, le risposte di difesa, le interazioni sociali e, in molti animali, la
riproduzione.
GUSTO
Il sistema gustativo riconosce le sostanze saporite presenti negli alimenti, che sono
soprattutto molecole idrosolubili o liposolubili. Queste molecole forniscono informazioni
sulla qualità, la quantità e l’eventuale pericolosità del cibo ingerito.
SISTEMA CHEMIOSENSORIALE DEL TRIGEMINO
Il sistema chemiosensoriale del trigemino fornisce informazioni sulle sostanze chimiche
irritanti o nocive che vengono a contatto con la pelle o le mucose degli occhi, del naso e
della bocca.
OLFATTO
Da un punto di vista evolutivo, i sensi chimici - in particolare l'olfatto - sono considerati i
sistemi sensoriali “più antichi”; tuttavia, rimangono per molti modi i meno compresi delle
modalità sensoriali.
Il sistema olfattivo elabora informazioni sull'identità, la concentrazione e la qualità di
un'ampia gamma di stimoli chimici volatili trasportati dall’aria, chiamati sostanze odorose.
Questi interagiscono con i neuroni olfattivi situati in un foglio epiteliale - l'epitelio olfattivo -
che riveste la superficie interna del naso.
ORGANIZZAZIONE DEL SISTEMA OLFATTIVO
I recettori olfattivi inviano i loro assoni direttamente
ai neuroni del bulbo olfattivo che, a sua volta, invia
proiezioni alla corteccia piriforme nel lobo temporale,
oltre che ad altre strutture del prosencefalo tramite
un fascio di assoni chiamato tratto olfattivo.
Il sistema olfattivo è unico tra i sistemi sensoriali
perché non comprende una stazione talamica,
interposta tra i recettori primari e la regione corticale
deputata all’elaborazione dell’informazione
sensoriale.
PECULIARITÀ DEL SISTEMA OLFATTIVO
All’interno del nostro naso sono presenti dei recettori che riconoscono le molecole presenti
nell’ara dell’odore della determinata osa a cui prestiamo attenzione. Questi recettori
riconoscono queste molecole e i trovano su dei neuroni che si chiamano neuroni olfattivi,
ch si trovano a loro volta incastonati nell'epitelio olfattivo - in questo caso, sostanze
chimiche odorose presenti nell'aria – interagiscono coi i recettori in periferia e vengono
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
trasdotte e codificate in segnali elettrici, che vengono quindi inoltrati a centri di ordine
superiore.
Meno si sa sull'organizzazione centrale del sistema olfattivo rispetto ad altri percorsi
sensoriali.
Non è ancora noto se nel bulbo olfattivo o nella corteccia piriforme esistano mappe
analoghe di specifiche sostanze odorose. I neuroni olfattivi vengono sostituti ogni 1-2 mesi.
vengono sostituiti graie alle cellule staminali basali chele rigenerano.
In effetti, fino a poco tempo fa è stato difficile immaginare quali qualità sensoriali sarebbero
state rappresentate in una mappa olfattiva o quali caratteristiche potrebbero essere
elaborate in parallelo come accade in altri sistemi sensoriali.
PERCEZIONE OLFATTIVA NELL’UOMO
Un gatto di 3 kg ha circa 20 cm2 di epitelio olfattivo. Nell'uomo, l'olfatto è spesso
considerato il meno sviluppato dei sensi e molti animali possiedono una capacità olfattiva
superiore a quella umana.
In un essere umano di 70 kg, la superficie dell'epitelio olfattivo è di circa 10 cm2.
Questa differenza può riflettere il maggior numero di recettori olfattivi e molecole recettrici
per sostanze odorose nell'epitelio olfattivo di altre specie e l'area proporzionalmente più
grande del prosencefalo dedicata all'olfatto. Allo stesso modo, la dimensione relativa del
bulbo olfattivo e delle strutture correlate rispetto agli emisferi corticali in un roditore o di un
carnivoro è piuttosto grande rispetto a quella nell'uomo.
LE CONCENTRAZIONI SOGLIA PER IL RILEVAMENTO E L'IDENTIFICAZIONE DELLE
VARIE MOLECOLE VARIANO NOTEVOLMENTE
L’uomo è in grado di identificare numerose sostanze odorose presenti nell’aria con un
ampio raggio e di identificarle come piacevoli, sgradevoli, irritanti, attraenti.
Differenze di concentrazione possono portare a differenze di percezione: l'indolo a bassa
concentrazione profuma di fiori, ad alte concentrazioni profuma di putrido.
Piccoli cambiamenti nella struttura molecolare possono anche portare a grandi differenze
percettive.
STRUTTURA CHIMICA E SOGLIA PERCETTIVA UMANA PER 12 ODORANTI COMUNI
Poiché il numero di odori è molto elevato, ci sono stati diversi tentativi di classificarli in
gruppi. Gli odori vennero divisi in categorie in base alla qualità percepita, alla struttura
molecolare e al fatto che alcune persone, chiamate anosmici, hanno difficoltà a annusare
l'uno o l'altro gruppo. Amoore classificava gli odoranti come pungenti, floreali, muschiati,
terrosi, eterei, canfora, menta piperita, putrido.
Tuttavia, questa classificazione rimane del tutto empirica. Gli odori più comuni in natura
sono miscele di diverse molecole odorose, anche se in genere sono vissute come un
singolo odore (come le percezioni suscitate dal profumo o dal «bouquet di un vino»).
Le molecole percepite a basse concentrazioni sono più liposolubili, mentre quelle con
soglie più elevate sono più solubili in acqua.
VALUTAZIONE DELLA FUNZIONE OLFATTIVA
Per “olfattometria” si intende un esame diagnostico sensoriale che consiste nell’impiego di
un kit per la presentazione standardizzata degli odoranti, a vari livelli di concentrazione, e
nella registrazione ed elaborazione statistica delle loro risposte, per ottenere il risultato
finale della misura. Fondamentalmente il paziente annusa specifici odori e si rileva la sua
capacità di rilevarli.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· iposmia, ovvero parziale riduzione del senso dell’odorato
· anosmia, ovvero totale riduzione del senso dell’odorato
· parosmia, un disturbo caratterizzato dall’alterazione del senso dell’olfatto
· cacosmia, cioè la percezione di odore sgradevole (allucinazioni olfattive)
Quando ai soggetti vengono presentati sette odori comuni, la stragrande maggioranza degli
individui "normali" è in grado di identificare correttamente tutti e sette gli odori (in questo
caso, borotalco, cioccolato, cannella, caffè, naftalina, burro di arachidi, e sapone). Alcune
persone, tuttavia, hanno difficoltà a identificare anche questi odori comuni.
PERDITA DELLA SENSIBILITÀ OLFATTIVA
La sensibilità e la percezione olfattiva può variare, oltre che per l’età, in diverse patologie e
disturbi tra cui:
· disturbi alimentari
· disturbi psicotici (in particolare la schizofrenia)
· diabete
· assunzione di determinati farmaci
· malattia di Alzheimer
tutti per ragioni che rimangono oscure
La capacità di identificare 80 odoranti comuni diminuisce notevolmente tra i 20 ei 70 anni.
Attivazione massima della corteccia orbitale frontale e della corteccia mediale cerebrale
(piriforme e amigdala) indotta da odoranti familiari in giovani adulti e in soggetti sani e
anziani.
LE SOSTANZE ODORANTI RISVEGLIANO COMPORTAMENTI INNATI CARATTERISTICI
In molte specie animali il sistema olfattivo mette in evidenza non solo gli odori ma anche i
feromoni, che sono sostanze liberate dagli animali che influenzano lo stato fisiologico di
altri membri della stessa specie.
Si ritiene che i feromoni attivino i neuroni sensoriali vomeronasali innescando risposte
sessuali, parentali o aggressive.
Nell’uomo non sono presenti organi vomeronasali e nonostante i tentativi di identificare
feromoni attivi, i dati a questo proposito sono modesti. Ciò nonostante le sostanze odorose
possono provocare varie risposte fisiologiche:
· Risposte motorie e viscerali all’aroma di cibi appetitosi (salivazione, aumento
motilità gastrica)
· Costrizione faringea, vomito, in presenza di cibi nocivi
· Possono influenzare la funzione endocrina e quella riproduttiva (ciclo mestruale
sincronizzati), oppure influenzare i rapporti tra madre e figlio (neonati che
riconoscono odore della madre e viceversa)
DIVERSA ATTIVAZIONE DELL’IPOTALAMO
L'esposizione ad una miscela di sostanze odorose, di androgeni ed estrogeni a
concentrazioni al di sotto del livello di rilevazione cosciente, possono suscitare sia risposte
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
comportamentali che differenti modelli di attivazione cerebrale in soggetti adulti di sesso
maschile e femminile.
Pertanto, sebbene la maggior parte degli umani non elabori i feromoni dal sistema
vomeronasale, altre strutture olfattive possono evidentemente rilevare segnali che possono
influenzare i comportamenti riproduttivi e altri comportamenti.
L’EPITELIO OLFATTIVO E I RECETTORI OLFATTIVI (v.slide 16)
Mucosa nasale
Sulla superficie apicale ogni recettore emette un unico processo dendritico che si espande
in una protuberanza da cui si estendono molti microvilli , ciglia olfattive, che si immergono
in uno stato di muco.
ALTRE FUNZIONI DELL’EPITELIO OLFATTIVO
La posizione superficiale della mucosa nasale consente ai recettori olfattivi di accedere
direttamente alle molecole odorose. A causa di questo i recettori sono esposti a inquinanti
dispersi nell'aria, allergeni, microrganismi e altre sostanze potenzialmente dannose che
sottopongono i recettori olfattivo a danni più o meno continui.
Numerosi meccanismi aiutano a mantenere l'integrità dell'epitelio olfattivo:
· Muco secreto dalle ghiandole di Bowman intrappola e neutralizza gli agenti
potenzialmente dannosi
· Le immunoglobuline, presenti nell'epitelio respiratorio che olfattivo, vengono
secrete nel muco, fornendo una difesa iniziale contro gli antigeni dannosi
· Le cellule di sostegno contengono enzimi (citocromo P450 e altri) che
catabolizzano composti organici potenzialmente dannosi
· Presenza di macrofagi
· I neuroni del recettore olfattivo sono sostituiti da un normale ciclo di
degenerazione e rigenerazione
RIGENERAZIONE DEI RECETTORI OLFATTIVI
Nei roditori, l'intera popolazione di neuroni olfattivi viene rinnovata ogni 6-8 settimane. La
rigenerazione si realizza mantenendo tra le cellule basali dell’epitelio olfattivo una
popolazione di precursori (cellule staminali) che si dividono per dare origine a nuovi
recettori sensoriali.
Questa rigenerazione spontanea delle cellule del recettore olfattivo offre l'opportunità di
studiare in che modo i precursori delle cellule nervose danno origine con successo a nuovi
neuroni che ricostituiscono connessioni sinaptiche e funzioni nel sistema nervoso centrale
dell’adulto, un argomento di ampio interesse clinico (neurogenesi).
I potenziali di recettore sono generati nelle ciglia dei neuroni olfattivi.
FAMIGLIA DI GENI DEI RECETTORI PER LE SOSTANZE ODOROSE
In tutti i mammiferi, i recettori per le sostanze odorose costituiscono la famiglia di geni più
numerosa tra quelle note e rappresentano il 2-5% di tutti i geni presenti nei mammiferi.
Il numero dei geni codificanti proteine funzionanti è minore per la presenza di pseudogeni.
Ogni sostanza viene riconosciuta da una particolare combinazione di recettori.
MECCANISMI MOLECOLARI DELLA TRASDUZIONE DEI SEGNALI OLFATTIVI
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
i neuroni olfattivi sono dei neuroni bipolari e alla loro estremità sono presenti delle ciglia.
Sulle ciglia vengono espressi dei recettori accoppiati a proteina G dove si lega la sostanza
odorante; una volta che il ligando si lega al recettore esso avvia una cascata di trasduzione
che coinvolge la proteina g che attiva la denilatociclasi. Si produce CMP che apre i canali
cationici che fanno entrare il Na e Ca = depolarizzazione.
1) La molecola odorosa si lega al recettore
2) G specifica per l’olfatto si dissocia
3) la subunità α si dissocia e attiva
4) l’ adenilato ciclasi III
5) aumenta AMP-ciclico
6) apertura dei canali nucleotide ciclico-dipendenti
7) ingresso ioni Na⁺ Ca²⁺
8) depolarizzazione
9) l'incremento della concentrazione di Ca²⁺ determina l’apertura di canali per il Cl che
contribuisce alla depolarizzazione del neurone
BULBO OLFATTIVO
I recettori olfattivi trasmettono tramite i loro assoni, i potenziali d'azione con le informazioni
relative alle sostanze odorose direttamente al cervello. Appena gli assoni lasciano l’epitelio
olfattivo si uniscono a formare un elevato numero di fasci che nel loro insieme costituiscono
il nervo olfattivo (I nervo cranico). Ogni nervo olfattivo proietta omolateralmente nel bulbo
olfattivo: struttura pari, di forma ovale, localizzata sotto la superficie ventrale del lobo
frontale.
ORGANIZZAZIONE DEL BULBO OLFATTIVO
I neuroni sensoriali che esprimono lo stesso tipo di recettore olfattivo convergono nello
stesso glomerulo nel bulbo olfattivo, dove formano sinapsi con i neuroni secondari
principali: le cellule mitrali (M) e le cellule a pennacchio (P), le quali proiettano
successivamente alla corteccia olfattiva. Gli interneuroni periglomerulari (PG) e le cellule
granulari (Gr), mediante l'inibizione laterale, concorrono a elaborare ulteriormente
l'informazione sensoriale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Quando il bulbo viene visto dalla sua superficie dorsale , si possono vedere glomeruli
olfattivi. Il denso accumulo di dendriti e sinapsi che costituiscono i glomeruli sono colorati
qui con un colorante fluorescente vitale che riconosce i processi neuronali. (B) Tra i
principali componenti neuronali di ciascun glomerulo vi sono i ciuffi apicali delle cellule
mitraliche, che si proiettano sulla corteccia piriforme e su altri target del bulbo. (C) La
struttura cellulare del bulbo olfattivo: sono indicati i vari strati del bulbo.
GLOMERULI
Sono strutture sferiche dove avvengono le sinapsi eccitatorie (glutammato) tra le afferenze
del recettore olfattivo e le cellule mitrali.
In ogni glomerulo arrivano informazioni da circa > 20.000 neuroni recettoriali olfattivi (25
cellule mitrali).
Interazioni inibitorie:
· all’interno dei glomeruli
· tra glomeruli distinti
Responsabile dell’attivazione selettiva di pochi glomeruli dominanti in presenza di odoranti
complessi: circuiti locali di inibizione laterale.
Ruolo nella plasticità sinaptica.
Coinvolto nella rigenerazione neuronale (forse).
Gli assoni dei recettori olfattivi che esprimono un determinato gene per una proteina
recettrice convergono su un piccolo sottogruppo di glomeruli bilateralmente simmetrici.
ORGANIZZAZIONE SCHEMATICA DEL SISTEMA OLFATTIVO
L'informazione sulle molecole odorose viene trasmessa dagli assoni delle cellule mitrali e
delle cellule a pennacchio del bulbo olfattivo, attraverso il tratto olfattivo, alla corteccia
olfattiva, composta da: nucleo olfattivo anteriore, tubercolo olfattivo, corteccia piriforme,
corteccia entorinale e amigdala.
Il nucleo olfattivo anteriore proietta alla corteccia piriforme sia ipsi- che controlaterale ma
anche al bulbo olfattivo.
Il tubercolo olfattivo è l'unica regione della corteccia olfattiva che non ha connessioni
efferenti al bulbo olfattivo e proietta principalmente al nucleo dorsomediale del talamo.
La corteccia piriforme, che riceve il maggior numero di proiezioni del bulbo, ha connessioni
dirette con la corteccia orbitofrontale, ma anche con il nucleo dorsomediale del talamo.
La corteccia entorinale proietta principalmente all'ippocampo ma anche al bulbo olfattivo,
mentre l'amigdala proietta sia all'ipotalamo che, nuovamente, al bulbo olfattivo.
Tutte le regioni della corteccia olfattiva primaria sono collegate da una complessa rete di
connessioni intracorticali.
Il bersaglio principale del tratto olfattivo è la corteccia piriforme a tre strati situata sul
versante ventromediale del lobo temporale, vicino al chiasma ottico.
I neuroni nella corteccia piriforme rispondono agli odori e le afferenze dalle cellule
mitraliche dei glomeruli, che ricevono proiezioni specifiche dai recettori olfattivi, sono
distribuite attraverso la corteccia piriforme.
Gli assoni delle cellule mitraliche formano un fascio - il tratto olfattivo laterale - che proietta
verso i nuclei olfattivi accessori, il tubercolo olfattivo, alla corteccia entorinale ed alcune
regioni dell’amigdala.
La corteccia piriforme ha dei neuroni piramidali che proiettano a una serie di bersagli nel
prosencefalo: diversi nuclei talamici, ipotalamici e verso l'ippocampo e l'amigdala.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Alcuni neuroni della corteccia piriforme innervano anche una regione della corteccia
orbitofrontale dove vengono prodotte risposte multimodali a stimoli complessi che
comprendono risposte a stimoli olfattivi e gustativi. Concetto di sapore: piacere nel
consumare cibi, valore edonico (gradevole o sgradevole) Comportamento alimentare
Emozioni: memoria olfattiva; formazione di nuova memoria dichiarativa relativa a un odore;
memoria evocata da un odore.
Le informazioni sugli odori raggiungono così una varietà di regioni del cervello,
permettendo ai segnali olfattivi di influenzare i comportamenti cognitivi, viscerali, emotivi e
omeostatici.
ORGANIZZAZIONE DELLA CORTECCIA PIRIFORME
I neuroni piramidali superficiali (PS), situati nello strato II, e i neuroni piramidali profondi
(PP), situati nello strato III, ricevono le afferenze dal bulbo olfattivo, che decorrono nello
strato I. A questo livello si ha anche l'integrazione mediata dai collaterali assonici dei
neuroni piramidali. Inoltre, nello strato III sono presenti varie classi di interneuroni inibitori
(I) che contribuiscono all'elaborazione dell'informazione sensoriale.
Non è stata descritta una mappa corticale spaziale dell’ambiente olfattivo: una sostanza
odorante attiva uno spettro ampio di neuroni nella corteccia olfattiva.
COMPONENTI CENTRALI DEL SISTEMA OLFATTIVO
I substrati neurali per l'elaborazione degli odori nell'uomo includono tutte le strutture
identificate anatomicamente come parte del percorso olfattivo: bulbo olfattivo, le cortecce
piriforme e orbitofrontali, amigdala; sono tutti chiaramente attivati dalla presentazione di
stimoli olfattivi nelle immagini fMRI di soggetti umani normali.
GUSTO
La funzione principale del gusto è di tipo nutritivo ed agisce di concerto con i sistemi
olfattivo e trigemino, indica se il cibo deve essere ingerito.
Le cellule del gusto trasducono questi stimoli e forniscono ulteriori informazioni sull'identità,
la concentrazione e la qualità piacevole o spiacevole della sostanza. Questa informazione
prepara anche il sistema gastrointestinale a ricevere cibo causando salivazione e
deglutizione (o la costrizione riflessa della faringe e il vomito se le sostanze sono nocive).
Le informazioni sulla temperatura e la consistenza del cibo (viscosità e contenuto di grassi)
vengono trasdotte ad opera dei recettori somatosensoriali del trigemino e di altri nervi
cranici e tramite questi nervi vengono trasmessi dalla lingua e dalla bocca al talamo e alle
cortecce somatosensoriali.
Una volta in bocca, i componenti chimici del cibo interagiscono con i recettori delle cellule
gustative situate in strutture specializzate dell’ epitelio linguale chiamate bottoni gustativi.
Il gusto dipende anche da fattori culturali e psicologici. Il cibo non viene semplicemente
consumato per valore nutritivo.
In quale altro modo si può spiegare perché a così tante persone piace consumare sostanze
piccanti o bevande dal sapore amaro?
PAPILLE, GEMME E CELLULE GUSTATIVE
Le papille filiformi non sono papille gustative. b, Morfologia delle papille circumvallate,
foliate e fungiformi contenenti gemme gustative. c, Ogni gemma gustativa è costituita da
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
varie cellule sensoriali gustative, ognuna delle quali è specializzata nella mediazione di un
tipo di sapore. La superficie apicale delle cellule gustative forma dei microvilli che entrano
in contatto, attraverso il poro gustativo, con gli stimoli chimici. All'estremità basale le cellule
comunicano con i neuroni gustativi afferenti.
In base ad un accordo generale tra diverse culture, esistono cinque categorie di gusto
percettivamente distinte: salato, acido, dolce, umami (dalla parola giapponese delizioso,
umami si riferisce a sapori salati, tra cui il glutammato monosodico e altri aminoacidi) e
amaro.
La percezione del gusto è strettamente legata alla biologia molecolare della trasduzione del
gusto.
Ogni parte della lingua è sensibile a ogni sapore, tuttavia parti diverse della lingua hanno
soglie diverse ai diversi sapori.
Diverse regioni della lingua hanno soglie diverse.
La punta della lingua è più sensibile ai composti dal sapore dolce e poiché questi composti
producono sensazioni piacevoli, le informazioni provenienti da questa regione attivano
comportamenti relativi all’alimentazione come movimenti della bocca, secrezione salivare,
rilascio di insulina e deglutizione.
Le risposte ai composti amari sono massime sulla parte posteriore della lingua.
L'attivazione di questa regione da parte di sostanze dal sapore amaro provoca protrusione
della lingua e altre reazioni protettive che impediscono l'ingestione. I composti dal sapore
acido suscitano smorfie, risposte increspate e un'enorme secrezione salivare per diluire il
gusto.
STIMOLI GUSTATIVI
La maggior parte degli stimoli gustativi sono molecole non volatili, idrofile solubili nella
saliva.
Alcuni esempi sono:
· sali come NaCl, necessari per l'equilibrio elettrolitico [10 mM]
· aminoacidi essenziali come il glutammato, necessari per la sintesi proteica
· zuccheri come il glucosio, necessari per produrre energia [20 mM]
· acidi come l'acido citrico che indicano l'appetibilità di vari alimenti (arance, nel
caso del citrato) [2 mM]
· molecole dal sapore amaro (compresi alcaloidi vegetali come atropina, chinino e
stricnina), che indicano alimenti che potrebbero essere velenosi
TRASDUZIONE DEL SEGNALE
Le cellule del gusto sono cellule epiteliali polarizzate con un dominio apicale e un dominio
basolaterale separati da giunzioni strette.
I canali di trasduzione degli stimoli gustativi (salato e
acido) e i recettori accoppiati a proteine G (dolce,
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
aminoacido e amaro) si trovano nel dominio apicale. Anche i componenti per la trasduzione
del segnale intracellulare (proteine G e secondi messaggeri) accoppiati alle proteine
recettrici degli stimoli gustativi si trovano nel dominio apicale.
I canali Na⁺, K⁺ e Ca²⁺ mediano il rilascio del neurotrasmettitore da strutture presinaptiche
specializzate che si trovano a livello basale sulle terminazioni di afferenze sensoriali
periferiche, così come il reticolo endoplasmatico che modula la concentrazione
intracellulare di Ca²⁺ contribuendo al rilascio di neurotrasmettitore.
Il neurotrasmettitore serotonina, insieme ad altri neurotrasmettitori e recettori della
serotonina, sono situati sulle terminazioni delle afferenze sensoriali. Infine, il canale
TRPM5, * che facilita la depolarizzazione mediata da recettori accoppiati a proteine G.
La sua localizzazione (nel dominio apicale o basale) non è ancora nota. * Potenziale del
recettore transitorio: 30 sottotipi nell'uomo.
Recettori per sali e acidi:
· Canali ionici
· Depolarizzazione e apertura canali per cationi
· Potenziale di recettore graduato
· Trasmissione sinaptica Ca2+-dipendente
· Rilascio serotonina (ATP o GABA)
· Potenziale di azione nel neurone afferente
Recettori per molecole dolci, amare e umami:
· Famiglia di 7 domini transmembrana accoppiata al sistema delle proteine G
· Sistema dei 2° messaggeri
· Vari effetti tra cui potenziale di recettore graduato e trasmissione sinaptica Ca2+
dipendente
· Rilascio serotonina (ATP o GABA)
· Potenziale di azione nel neurone afferente
MECCANISMI MOLECOLARI
La selettività cationica di Na+ sensibile all'amiloride rispetto al canale protonico sensibile
all'H+ fornisce la base per la specificità dei sapori salati e acidi. In ogni caso, la corrente
positiva attraverso il canale cationico porta alla depolarizzazione della cellula. (PDK:
malattia renale policistica).
Per i sapori dolci, aminoacidi (umami) e amari, diverse classi di recettori accoppiati alle
proteine G mediano la trasduzione. Per il dolce, i complessi eterodimerici dei recettori
T1R2 e T1R3 trasducono gli stimoli attraverso un meccanismo mediato da PLCᵦ₂ ,
dipendente da IP3 che porta all'attivazione del canale TRPM5per il Ca²⁺(PLCᵦ₂ : isoforma
della fosfolipasi C). Per gli aminoacidi, i complessi eterodimerici dei recettori T1R1 e T1R3
trasducono gli stimoli attraverso lo stesso meccanismo dipendente da PLCᵦ₂ / IP₃ / TRPM5
I sapori amari vengono trasdotti attraverso un insieme distinto di recettori accoppiati alle
proteine G, i sottotipi del recettore T2R (ad oggi sono noti 30 sottotipi T2R). I recettori T2R
sembrano associati alla proteina G specifica per le cellule gustative: gustoducina che non
si trova nelle cellule del gusto che esprimono i recettori degli aminoacidi o dolci. Tuttavia, la
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
depolarizzazione accoppiata allo stimolo per i gusti amari si basa sullo stesso meccanismo
PLCᵦ₂ / IP₃ / TRPM5 dipendente usato per la trasduzione del gusto dolce e aminoacidico.
RECETTORI E MECCANISMI DI TRASDUZIONE DEL GUSTO
Molecole dal sapore amaro, dolce o umami attivano specifiche combinazioni di recettori (R)
delle famiglie T1R e T2R che innescano una cascata di trasduzione metabotrobica.
L'attivazione dei recettori porta all'attivazione della fosfolipasi PLCᵦ₂ mediata dalla
gustoducina. La produzione di inositolo trisfosfato (IP3) determina la liberazione di calcio
dal reticolo endoplasmatico che attiva il canale TRPM5, provocando la depolarizzazione
della cellula.
Il sapore salato è principalmente dovuto a ioni sodio che attraversano canali per il sodio
sensibili all'amiloride (ENaC).
Il sapore acido è mediato principalmente dai canali ionici PKD2L1 e PKD2L3.
Meccanismi di comunicazione all'interno delle gemme gustative. Le cellule di tipo II, attivate
da stimoli gustativi, liberano adenosin-trifosfato (ATP) che agisce sui neuroni afferenti.
L'ATP induce anche la liberazione di 5HT da parte delle cellule di tipo III, che agisce sia
sulle fibre afferenti sia sulle cellule di tipo II.
ELABORAZIONE DEL GUSTO A LIVELLO DEL SISTEMA NERVOSO CENTRALE
Il sistema gustativo comprende sia i recettori periferici che una serie di vie centrali. Le
cellule del gusto (i recettori periferici) si trovano nelle papille gustative distribuite sulla
superficie dorsale della lingua, sul palato molle, sulla faringe e sulla parte superiore
dell'esofago. Queste cellule formano sinapsi con assoni sensoriali primari che corrono nella
corda del timpano e nelle ramificazioni del gran petroso superficiale del nervo facciale
(nervo cranico VII), nel ramo linguale del nervo glossofaringeo (nervo cranico IX) e il ramo
laringeo superiore del vago nervo (nervo cranico X) per innervare i bottoni gustative
rispettivamente della lingua, del palato, dell'epiglottide e dell'esofago.
Gli assoni centrali di questi neuroni sensoriali primari nei rispettivi gangli dei nervi cranici
proiettano verso le regioni rostrale e laterale del nucleo del tratto solitario nel midollo
allungato, detto anche come nucleo gustativo del tratto solitario. Gli assoni dalla parte
rostrale (gustativa) del nucleo solitario proiettano al complesso ventrale posteriore del
talamo, dove terminano nella porzione mediale del nucleo ventrale posteriore mediale.
Questo nucleo si proietta a sua volta verso diverse regioni della corteccia, tra cui l'insula
anteriore nel lobo (corteccia gustativa insulare) temporale e l'opercolo del lobo frontale
(area 43).
Esiste anche un'area di gusto corticale secondaria nella corteccia orbitofrontale nella parte
caudale, in cui i neuroni rispondono alle combinazioni di stimoli visivi, somatosensoriali,
olfattivi e gustativi.
Le proiezioni reciproche che collegano il nucleo del tratto solitario attraverso il ponte
all'ipotalamo e all'amigdala, presumibilmente influenzano aspetti affettivi dell'appetito, della
sazietà e altre risposte omeostatiche associate all'alimentazione.
La parte caudale del nucleo del tratto solitario riceve anche innervazione da rami
subdiaframmatici del nervo vago, che controllano la motilità gastrica. Gli interneuroni che
collegano le regioni rostrale e caudale del nucleo rappresentano la prima interazione tra
stimoli viscerali e gustativi e questo circuito può essere considerato la componente
sensoriale di un arco riflesso gustativo-viscerale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Questa stretta relazione di informazioni gustative e viscerali ha un preciso significato, dal
momento che un animale deve riconoscere rapidamente se sta mangiando qualcosa che
potrebbe fargli male e risponde di conseguenza.
È interessante notare che quando un determinato cibo viene consumato al punto di
sazietà, specifici neuroni orbitofrontali nella scimmia riducono la loro attività a quel gusto,
suggerendo che questi neuroni sono coinvolti nella motivazione consapevole a mangiare (o
non mangiare) cibi particolari.
IL SISTEMA CHEMIOSENSORIALE DEL TRIGEMINO
Il sistema chemiosensoriale del trigemino è costituito da neuroni nocicettivi polimodali e dai
loro assoni nel nervo trigemino (nervo cranico V) e, in misura minore, neuroni nocicettivi i
cui assoni corrono nei nervi glossofaringeo e vago (IX e X).
Questi neuroni e le loro terminazioni associate sono in genere attivati da sostanze chimiche
classificate come irritanti, inclusi inquinanti atmosferici (ad es. Anidride solforosa),
ammoniaca (sali odoranti), etanolo (liquore), acido acetico (aceto), anidride carbonica
(nelle bevande analcoliche), mentolo (in vari inalanti) e capsaicina (il composto presente
nei peperoncini piccanti che suscita la caratteristica sensazione di bruciore) o nocive che
entrano in contatto con pelle o mucose occhi, naso, bocca.
I nocicettori polimodali per le sostanze irritanti avvisano l'organismo di stimoli chimici
potenzialmente dannosi che sono stati ingeriti, inspirati o che entrano in contatto con il viso.
LA CAPSAICINA
Se applicata alla mucosa orale agisce da irritante innescando reazioni di protezione:
lacrimazione, bruciore, sudorazione.
Una varietà di risposte fisiologiche mediate dal sistema chemiosensoriale trigemino sono
innescate dall'esposizione a sostanze irritanti. Questi includono:
· aumento della salivazione
· vasodilatazione
· lacrimazione
· secrezione nasale
· sudorazione
· broncocostrizione
I recettori delle sostanze irritanti si trovano principalmente sui rami terminali dei neuroni
nocicettivi polimodali. Sebbene questi recettori rispondano a molti degli stessi stimoli dei
neuroni del recettore olfattivo (ad es. aldeidi, alcoli), probabilmente non sono attivati dallo
stesso meccanismo (i recettori accoppiati alle proteine G per gli odori si trovano solo nei
neuroni del recettore olfattivo).
Ad eccezione della capsaicina e degli stimoli acidi, che attivano entrambi i canali ionici
selettivi per cationi; si sa poco sui meccanismi di trasduzione degli irritanti o sulla loro
elaborazione centrale.
Nocicettori polimodali: possono essere attivati da stimoli meccanici, chimici, o termici (caldo
o freddo intenso).
SISTEMA SOMATOSENSORIALE
cellula recettoriale: elemento chiave di tutte le modalità sensoriali
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
il recettore è l’interfaccia tra il sistema nervoso e il mondo esterno → trasforma gli stimoli
provenienti dl mondo esterno in qualcosa in cui il sistema nervoso è in grado di elaborare.
- questi recettori hanno la caratteristica di essere specializzati per una data modalità
sensoriale
>recettori specializzati per gli stimoli luminosi, per le vibrazioni…
- all’interno della modalità sensoriale per cui questi recettori sono specifici, troviamo
che ci sono delle sottotipologie di recettori che sono particolarmente sensibili per
certi ambiti.
Recettori sensoriali:
· stimoli ai quali possono rispondere
· vie sensoriali che ritrasmettono informazioni dai recettori alla corteccia cerebrale
Ogni modalità sensoriale è mediata da un sistema neurale diverso che è composto di
diverse componenti che contribuiscono alla percezione. I sistemi sensoriali sono costituiti
da neuroni che mettono in connessione i recettori periferici con il midollo spinale, il tronco
dell’encefalo, il talamo e la corteccia cerebrale.
INFORMAZIONE SENSORIALE
Informazione sensoriale: attività nervosa che prende origine dalla stimolazione di cellule
recettoriali disposte in particolari sedi corporee. Le sensazioni destate da questi recettori
comprendono i 5 sensi e altre modalità sensoriali che non erano riconosciute nell’antichità
ma che sono essenziali per le funzioni dell’organismo:
· Sensazioni somatiche della propriocezione (postura e movimento del proprio corpo)
dolore, prurito, temperatura corporea;
· Sensazioni viscerali (coscienti e inconsce) che sono necessarie per l’omeostasi;
· Sensazioni vestibolari dell’equilibrio : posizione del corpo all’interno del campo
gravitazionale e movimento del capo.
In ogni sistema sensoriale i recettori forniscono la prima rappresentazione nervosa del
mondo esterno.
SENSIBILITÀ
è il concetto per cui un recettore risponde maggiormente ad uno stimolo; se esso ricade
nella gamma per cui è più specifico.
Il sistema sensitivo comprende:
· Recettori: funzionano da trasduttori. Percepiscono stimoli provenienti
dall’ambiente esterno (esterocettori), da organi interni (enterocettori) e da
articolazioni e muscoli (propriocettori) e li trasformano in segnali interpretabili
dal SNC (potenziale d’azione).
· Neuroni sensitivi primari: in rapporto con i recettori inviano l’informazione
relativa alle caratteristiche dello stimolo ai centri nervosi attraverso le fibre
afferenti sensoriali.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Neuroni sensitivi secondari ricevono informazioni dai neuroni primari e le inviano
ai neuroni terziari (talamo) che le trasmettono alla zona di corteccia cerebrale
specifica per quel tipo di sensibilità.
Il raggiungimento del livello di coscienza di uno stimolo sensoriale prende il nome di
sensazione. Le informazioni sensoriali provenienti dagli organi interni e da articolazioni e
muscoli non raggiungono il livello di coscienza ma danno origine a risposte finalizzate al
mantenimento dell’omeostasi (afferenze viscerali) o al controllo del comportamento motorio
(afferenze propriocettive).
DIVISIONE SENSORIALE DEL SISTEMA NERVOSO
INFORMAZIONI CONSCE
Sensi speciali:
· Vista
· Udito
· Gusto
· Olfatto
· Equilibrio
Sensi somatici:
· Tatto-pressione
· Temperatura
· Dolore
· Propriocezione
INFORMAZIONI INCONSCE
Stimoli somatici:
· Lunghezza
· Tensione muscolare
Stimoli viscerali:
· Pressione arteriosa
· pH del liquido cerebrospinale
· Inflazione polmonare
· Temperatura
PROPRIETÀ GENERALI DEI SISTEMI SENSORIALI
I recettori sono strutture nervose (semplici o complesse) deputate a captare le diverse
forme di energia (stimoli) che agiscono sull’organismo (interno e/o esterno) e a trasformare
(trasduzione) queste ultime in segnali nervosi che vengono convogliati da apposite vie
nervose afferenti ai centri dove viene elaborata la risposta.
Stimolo → Recettore (via afferente) → Centro nervoso sensitivo
PRINCIPALI MODALITÀ SENSORIALI DELL’UOMO
· Vista
· Olfatto
· Gusto
· Tatto
· Sensi termici
· Dolore
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Udito
· Equilibrio
· Propriocezione
CLASSIFICAZIONE DEI RECETTORI:
· Posizione o origine dello stimolo
· Natura dello stimolo
· Rapporti con altre catene neuroniche afferenti
· Adattamento allo stimolo
Posizione o origine dello stimolo:
Meccanocettori -Tattili
-Labirintici acustici e vestibolari
-Fusi neuromuscolari, di Golgi e
articolari
Termocettori -Per il caldo
-Per il freddo
Chemiocettori -Olfattivi
- Gustativi
-Dei glomi aortici e carotidei
Nocicettori -Terminazioni libere superficiali o
profonde
Fotorecettori -Coni
-Bastoncelli
Natura dello stimolo:
Meccanocettori -Barocettori (pressione)
-Osmocettori (distensione cellulare),
vibrazione, accelerazione, onde
sonore, stiramento muscolare,
tensione muscolare
Termocettori -vari gradi di calore o di freddo
Fotorecettori -fotoni
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Chemiorecettori - ossigeno, pH, molecole organiche
come il glucosio
Volorecettori -volume liquido corporei
Caratteristiche morfologiche:
· Recettori semplici: terminazioni nervose libere con assoni mielinizzati o non
mielinizzati (es recettori dolorifici)
· Recettori complessi: una terminazione nervosa racchiusa in una capsula di
connettivo (es. Corpuscoli di Pacini) o differenziata (es. corpuscoli di Ruffini)
· Cellule recettoriali specializzate: fanno sinapsi su un neurone sensoriale. Es.
recettori uditivi, vestibolari, gustativi
Rapporti con altre catene neuroniche afferenti:
· Recettori di I tipo: terminazioni nervose libere o capsulate che hanno la loro
cellula (neurone di I ordine) nei gangli spinali o cerebrali (es. recettori cutanei,
mucosi, muscolari, olfattivi, ecc)
· Recettori di II tipo: cellule neuroepiteliali che sinaptano immediatamente con il
prolungamento periferico della cellula gangliare (neurone di I ordine). Es. cellule
labirintiche o gustative)
· Recettori di III tipo: cellule altamente differenziate in rapporto con una catena
neuronica multipla (es coni e bastoncelli retinici)
GLI ATTRIBUTI ELEMENTARI DELLO STIMOLO COMPONGONO LA SENSAZIONE
1. Modalità: definisce una classe generale di stimoli in base al tipo di energia che
gli stimoli trasmettono.
2. Sede: è rappresentata dall’insieme dei recettori di un sistema sensoriale che
vengono attivati. All’interno del sistema i recettori hanno una topografia e una
distribuzione spaziale ben definita che segnala la posizione dello stimolo.
3. Intensità: viene segnalata dall’ampiezza della risposta di ogni recettore, che è in
rapporto con l’ammontare complessivo dell’energia dello stimolo liberata a livello
del recettore stesso.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
4. Durata: viene definita dall’inizio e dalla fine della risposta del recettore e dipende
dalla velocità con cui lo stimolo comincia ad essere efficace e cessa di esserlo a
livello del recettore.
Tutti i sistemi sensoriali, nonostante la diversità delle sensazioni, quando vengono stimolati
ritrasmettono segnali concernenti i 4 attributi elementari degli stimoli.
IL SISTEMA SOMATOSENSITIVO
Le informazioni somatiche vengono fornite da recettori disposti in tutto il corpo.
funzioni principali:
· propriocezione: consapevolezza della postura e dei movimenti del nostro corpo, in
modo particolare dei quattro arti e del capo.
Recettori: delle capsule articolari / della cute /dei muscoli scheletrici
· esterocezione: concerne il senso di interazione diretta con il mondo esterno quando
entra in contatto con il corpo
· enterocezione: riguarda il senso delle condizioni funzionali dei principali sistemi
dell’organismo e del suo stato interno.
Alcune componenti del tatto richiedono una componente motoria attiva. Le componenti
sensitiva e motoria sono connesse dal punto di vista anatomico a livello del SNC e sono
importanti per la guida del comportamento. Comprende anche:
- i sensi termici del caldo e del freddo: importanti per mantenere il controllo del
comportamento e per mantenere la temperatura corporea vicino ai 37˚C.
- nocicezione:
risposta a eventi esterni che recano danno all’organismo. Meccanismo alla
base dei comportamenti motivati necessari per la sopravvivenza: fuga o combattimento.
Tutte le sensazioni somatiche sono mediate da una sola classe di neuroni sensitivi: i
neuroni dei gangli delle radici dorsali.
TATTO
Il tatto è quella modalità sensoriale rappresentata dalla meccanocezione superficiale, vale
a dire da recettori sensibili a stimoli meccanici che si trovano a livello cutaneo. La
meccanocezione comprende tre qualità: pressione (intensità e tempo), tatto in senso stretto
(velocità), vibrazione (accelerazione).
I meccanocettori sono connessi a fibre di tipo Aβ di grande diametro e alta velocità di
conduzione garantendo una rapida trasmissione delle informazioni al SNC. Le fibre Aβ non
sono altro che gli assoni periferici dei neuroni a T (neuroni sensitivi primari) dei gangli delle
radici dorsali.
MECCANOCETTORI
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Corpuscolo di Meissner: molto superficiali, meccanocettori della pelle glabra. Attivati
da pressioni anche molto leggere. Mostrano un aumento di scarica all’inizio della
stimolazione, seguita da una brusca riduzione dell’attività nervosa (RA1).
· Corpuscolo di Pacini: situati in profondità nei tessuti sottocutanei, nelle membrane
interossee e nell’intestino. Hanno una capsula a strati concentrici e sono a rapidissimo
adattamento. Senso di vibrazione e riconoscimento delle superfici degli oggetti.
Stimolazione selettiva delle fibre afferenti del Pacini determinano sensazione vibratoria e
solletico. Genera un solo potenziale d’azione all’inizio della stimolazione seguito da un
silenzio dell’attività nervosa (RA2).
· Disco di Merkel: superficiali, subito sotto l’epidermide. A lento adattamento (recettori
LA1) si trovano soprattutto su dita e labbra. Ruolo nella stimolazione statica della cute,
riconoscimento dei margini e delle forme degli oggetti poggiati sulla cute. Mostrano
un’alta frequenza di scarica all’inizio della stimolazione che diminuisce con il tempo.
· Corpuscolo di Ruffini: si trovano in profondità della cute, nei legamenti e nei tendini.
Hanno un lento adattamento (LA2) e loro forma allungata a fuso li rende adatti a
registrare la tensione prodotta dai movimenti delle dita e degli arti. Hanno un pattern di
scarica simile ai SAI: alta frequenza di stimolo all’inizio che diminuisce lentamente.
CLASSIFICAZIONE DELLE FIBRE SENSITIVE DEI NERVI PERIFERICI
Le diverse modalità della sensibilità somatica sono mediate da fibre nervose periferiche
che differiscono per il diametro e la velocità di conduzione.
NEURONI DEI GANGLI DELLE SUPERFICI DORSALI
I neuroni pseudounipolari hanno il soma posto di lato e un singolo processo che poi si
divide a T: un processo periferico che trasporta l’impulso dalla periferia al soma e un
processo centrale che trasporta un impulso dal soma al SNC.
Gli assoni hanno due rami, uno proietta alla periferia e l’altro al SNC. I rami periferici di
questi neuroni innervano i bersagli che stanno alla periferia, le loro terminazioni fungono da
recettori specializzati per particolari tipi di stimoli, che differiscono per morfologia e
selettività agli stimoli.
Ogni neurone DRG forma un’unica linea di trasmissione con un’unica polarità fra la
terminazione recettoriale e il SNC e viene detto fibra afferente primaria.
Le fibre afferenti primarie che innervano una particolare regione somatica si dispongono in
gruppi formando fasci o fascicoli di fibre denominati nervi periferici.
Lesioni dei nervi periferici o dei loro bersagli a livello del SNC provocano deficit sensitivi.
I rami centrali terminano nel midollo spinale o nel tronco dell’encefalo e formano la prima
sinapsi delle vie somatosensitive.
Le informazioni provenienti dalla cute, muscoli, articolazioni, visceri vengono ritrasmessi
dai neuroni dei gangli delle radici dorsali (DRG) che innervano gli arti e il tronco o dai
neuroni sensitivi del trigemino che innervano le regioni del capo.
SISTEMA AFFERENTE
1. Gli assoni afferenti primari portano l’informazione dai recettori sensoriali somatici
(distribuiti in tutto il corpo) al midollo spinale o al tronco encefalico.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
2. Gli assoni afferenti primari entrano nel midollo attraverso le vie dorsali, i loro corpi
cellulari si trovano nei gangli delle radici dorsali.
3. Attraverso il midollo spinale arrivano alla corteccia somatosensitiva.
IL DERMATOMERO
Tra l’organizzazione segmentale dei nervi spinali e l’innervazione sensoriale della pelle vi è
una correlazione. La regione cutanea innervata dalle radici dorsali di destra e di sinistra è
chiamata dermatomero, con corrispondenza univoca tra i dermatomeri e i segmenti spinali.
Quando la radice dorsale è tagliata, il dermatomero corrispondente non perde tutte le
sensazioni, perché le radici dorsali adiacenti innervano aree sovrapposte.
Si può mappare la pelle innervata dagli assoni della radice dorsale in caso di herpes zoster
(Fuoco di Sant’Antonio).
Se mappati,i dermatomeri disegnano una serie di zone sulla superficie corporea.
HERPES ZOSTER (FUOCO DI SANT’ANTONIO)
Dolori lancinanti per un’aumentata eccitabilità dei neuroni sensoriali, abbassando
notevolmente la soglia di scarica. Pelle infiammata e vescicolosa, ipersensibile a ogni
stimolo (iperestesia). Si riattiva solo in un ganglio della radice dorsale: i sintomi sono limitati
agli assoni della radice dorsale interessata e dalla pelle da essa innervata.
Il virus determina chiaramente sulla pelle il territorio di un dermatomero. Osservazioni su
questi pazienti sono state utili per mappare i dermatomeri.
LA VIA COLONNA DORSALE-LEMNISCO MEDIALE
Le fibre afferenti primarie che ritrasmettono informazioni tattili penetrano nella colonna
dorsale ipsilaterale dove ascendono al bulbo senza incrociarsi. Le fibre provenienti dalle
parti inferiori decorrono nel funicolo gracile e terminano nel nucleo gracile, mentre quelle
provenienti dalle parti superiori del corpo decorrono nel funicolo cuneato e terminano nel
nucleo cuneato.
Gli assoni dei due nuclei passano poi dal lato controlaterale del tronco encefalico e
ascendono al talamo formando un lungo fascio di fibre detto lemnisco mediale. Del
lemnisco mediale sono disposte in maniera somatotopica. Poiché queste fibre attraversano
la linea mediana per portarsi dall’altro lato, l’emisfero cerebrale destro riceve informazioni
sensitive dal lato sinistro del corpo e viceversa.
Le fibre del lemnisco mediale terminano a livello di una specifica suddivisione del talamo,
denominata nucleo ventrale posteriore.
Queste fibre mantengono una organizzazione somatotopica: le fibre che trasmettono
informazioni dalle parti inferiori del corpo terminano lateralmente, informazioni dalle parti
superiori e della faccia terminano medialmente.
LE VIE TRIGEMINALI
La sensazione somatica del volto, è dovuta alle tre branche trigeminali (oftalmica,
mascellare, mandibolare), che arriva al SNC a livello del ponte. È il V paio di nervi cranici,
innerva la faccia, l’area periorale e i due terzi più esterni della lingua.
Le connessioni sensoriali del nervo trigemino sono analoghe a quelle delle radici dorsali.
Gli assoni sensoriali a largo diametro del nervo trigemino recano informazioni tattili dai
meccanocettori cutanei. Essi fanno sinapsi nei neuroni di secondo ordine nel nucleo
trigeminale ipsilaterale, che analogo al nucleo della colonna dorsale. Gli assoni del nucleo
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
trigeminale decussano e proiettano nella parte ventrale postero mediale del talamo. Da qui
l’informazione è inviata alla corteccia somatosensoriale.
IL TALAMO
Di forma ovale nella parte dorsale del diencefalo, ritrasmette le informazioni sensoriali alle
aree sensoriali primarie della corteccia cerebrale.
Alcuni ricevono informazioni specifiche relative a una sola modalità sensoriale e proiettano
a una specifica area del neocortex: es nucleo ventrale postero laterale, a livello del quale
termina il lemnisco mediale, elaborano informazioni sensitive e i loro assoni proiettano alla
corteccia somatosensitiva primaria.
Altri nuclei prendono parte a funzioni motorie, ritrasmettendo informazioni dal cervelletto e
dai nuclei della base alle regioni motorie del lobo frontale.
La sua funzione non è semplicemente di centro di trasmissione ma filtra le informazione in
arrivo. Impedisce o potenzia il passaggio alla corteccia cerebrale di informazioni specifiche
a secondo dello stato comportamentale dell’animale.
Gli assoni delle cellule del talamo che proiettano alla neocorteccia decorrono nella capsula
interna: cospicuo fascio di fibre costituito per la maggior parte di fibre che lasciano o
raggiungono gli emisferi cerebrali.
Le connessioni con il lobo frontale lo portano ad avere un ruolo nei processi cognitivi es.
memoria, attenzione.
Il nuclei del talamo vengono classificati in 4 gruppi: anteriore, mediale, ventrolaterale e
posteriore; facendo riferimento alla lamina midollare interna, che è un fascio di fibre di
forma laminare che decorre per tutta l’estensione rostro-caudale del talamo.
Il nucleo reticolare (che forma una sorta di rivestimento esterno) non proietta alla corteccia
ma è costituito da neuroni inibitori. Questi ricevono afferenze dalle collaterali di fibre che
fuoriescono dal talamo, si portano al neocortex e proiettano a loro volta ad altri nuclei
talamici.
GRUPPO ANTERIORE E GRUPPO MEDIALE
Il gruppo anteriore riceve le sue principali afferenze dai nuclei mammillari dell’ipotalamo e dal
subiculum della formazione dell’ippocampo, è interconnesso con regioni della corteccia del
cingolo e del lobo frontale. Il ruolo funzionale del nucleo anteriore non è stato ancora
identificato, si ritiene essere implicato nella memoria e nelle emozioni.
Il gruppo mediale è costituito dal nucleo dorsomediale. Questo presenta tre suddivisioni,
ciascuna delle quali è connessa con una particolare regione della corteccia frontale. Esso
riceve afferenze dai nuclei della base, dall’amigdala ed è implicato nella memoria.
GRUPPO VENTRALE E GRUPPO POSTERIORE
I nuclei del gruppo ventrale sono denominati a seconda della loro posizione all’interno del
talamo. Quello ventrale anteriore e ventrale laterale sono importanti per il controllo motorio e
ritrasmettono informazioni dai nuclei della base e dal cervelletto alla corteccia motoria. Il
nucleo ventrale posterolaterale convoglia informazioni somatosensitive al neocortex.
Il gruppo posteriore comprende il corpo genicolato mediale e quello laterale, il nucleo
laterale posteriore e il pulvinar. Il corpo genicolato mediale e quello laterale sono situati nella
parte posteriore del talamo.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Nuclei aspecifici: proiettano a parecchie regioni corticali e sottocorticali e sono
quelli denominati della linea mediana. Si ritiene che controllino il livello di
vigilanza della corteccia.
· Nucleo reticolare: i suoi neuroni sono GABAergici e non sono interconnessi con
il neocortex ma i suoi assoni finiscono in altri nuclei talamici e forniscono
afferenze a questo nucleo che modula l’attività degli altri nuclei in base al flusso
di informazioni talamo-corticali.
· Nuclei di ritrasmissione (o specifici): stabiliscono relazioni specifiche con il
neocortex.
ORGANIZZAZIONE SOMATOTOPICA E DISTRIBUZIONE DEI CAMPI RECETTIVI DEI
NEURONI NEL NUCLEO VENTROPOSTEROLATERALE (VPL) E
VENTROPOSTEROMEDIALE (VPM) DEL TALAMO
VPM del talamo è rappresentata la faccia con una preponderanza delle labbra e della zona
peribuccale. Nel nucleo VPL è rappresentato il resto del corpo con preponderanza della
mano.
Rappresentazione significa che determinati neuroni ricevono informazioni da una parte
specifica del corpo: più questa è ricca di recettori, maggiore è la rappresentazione a livello
del SNC, cioè maggiore è il numero di neuroni che ricevono informazioni da questa parte
del corpo.
La rappresentazione del corpo nel SNC non dipende dalle reali dimensioni, ma dalla
densità di recettori in essa presenti.
Es: polpastrelli della mano hanno una rappresentazione enorme. Al contrario il torace ha
una superficie molto più grande di un polpastrello ma minore densità di recettori, quindi la
sua rappresentazione centrale è molto piccola.
CAMPI RECETTIVI
Esiste una relazione molto stretta tra rappresentazione centrale e campi recettivi. Maggiore
è la parte del corpo minore è la dimensione dei campi recettivi.
I polpastrelli hanno campi recettivi molto piccoli (pochi mm di diametro) mentre il torace ha
campi recettivi molto grandi (diversi cm).
Quelle parti del corpo che devono servire ad esplorare l’ambiente circostante devono avere
molti recettori e una grande rappresentazione nel cervello con neuroni con campi recettivi
piccoli per aumentare il contrasto degli oggetti esplorati.
L’acuità tattile è la capacità di distinguere due stimoli tattili, ad esempio due punte di
compasso separate l’una dall’altra.
CAMPI RECETTIVI DEI NEURONI SENSORIALI
Diversi neuroni sensitivi primari possono convergere e fare sinapsi su un singolo neurone
sensitivo secondario. I loro singoli campi recettivi pertanto si fondono per formare un unico
grande campo recettivo.
La convergenza dei neuroni sensoriali primari permette a stimoli simultanei sottosoglia di
sommarsi al livello del neurone secondario e innescare un potenziale d’azione.
INIBIZIONE LATERALE
L’attivazione di un neurone secondario più vicino alla sede in cui ha agito lo stimolo induce
due effetti:
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
1. eccita una via inibitoria che sopprime la risposta dei neuroni sensoriali periferici allo
stimolo
2. simultaneamente permette alla via sensitiva più vicina allo stimolo di procedere senza
interferenze
Es. Il neurone sensitivo secondario B, quando attivato, invia segnali inibitori alle adiacenti
vie A e C tramite collaterali. Questa inibizione diminuisce l’attività della via A e C e, per
contrasto, fa sembrare maggiore l’intensità dello stimolo a livello di B.
CORTECCIA SOMATOSENSITIVA PRIMARIA
I neuroni somatosensitivi di un lato del corpo proiettano al lato opposto del cervello. Lo
spazio della corteccia cerebrale destinato a ogni segmento corporeo è proporzionale alla
sensibilità di quel segmento. La sensibilità di un determinato segmento corporeo dipende
dalla densità dei recettori ivi presenti.
ORGANIZZAZIONE S-I (SOMATOSENSITIVA PRIMARIA)
La sproporzione dell’homunculus dipende dalla densità dei recettori.
I segnali che riceve S-I rappresentano almeno la terza stazione sinaptica.
I segnali sono stati elaborati da: colonne dorsali; talamo; corteccia stessa. Ogni neurone
corticale riceve segnali afferenti dai recettori periferici, in regioni specifiche della cute che
contribuiscono a definire il proprio campo recettivo. Percepiamo che una determinata sede
è stata toccata perché viene attivata nel SNC una specifica popolazione di neuroni. Stessa
esperienza se viene stimolata elettricamente la stessa popolazione di neuroni corticali.
La corteccia somatosensitiva primaria è formata da quattro aree citoarchitettoniche: 3a, 3b,
1 e 2 di Brodmann.
ELABORAZIONE DELLE INFORMAZIONI
Le aree citoarchitettoniche sono estensivamente interconnesse fra loro e l’elaborazione
dell’informazione avviene sia in serie che in parallelo.
Le informazioni somatosensitive vengono ritrasmesse in parallelo dalle 4 aree della S-I ai
centri di ordine superiore, che comprendono: corteccia S-II, corteccia parietale posteriore,
corteccia motoria.
Le aree 3b e 1 ricevono informazioni dai recettori cutanei, mentre 3a e 2 ricevono
informazioni di natura propriocettiva dai recettori dei muscoli, articolazioni, cute.
Le aree 3a e 3b ricevono afferenze dal talamo (nucleo ventrale posterolaterale, mediale,
superiore) e proiettano alle aree 1 e 2.
ORGANIZZAZIONE SOMATOTOPICA DELLA RAPPRESENTAZIONE CORPOREA
NELL'AREA SOMESTESICA PRIMARIA
Nella circonvoluzione postcentrale esistono quattro rappresentazioni della periferia
corporea, una per ognuna delle aree citoarchitettoniche (3a, 3b, 1 e 2) che compongono
l'area somestesica primaria.
Le quattro rappresentazioni della periferia seguono lo stesso piano organizzativo, con le
regioni caudali del corpo localizzate medialmente e quelle cefaliche lateralmente.
BARREL CORTEX
Le sviluppate vibrisse dei roditori meritano un ampia zona della S-I. I segnali sensoriali
provenienti da ciascun follicolo della vibrissa si dirigono versi definiti gruppi neuronali detti
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
«barrels». Le cinque file di vibrisse corticali mappano in modo preciso le cinque fila di
vibrisse facciali.
HOMUNCULUS PLASTICO
Le mappe sensoriali della superficie corporea presenti in corteccia non sono fisse, ma si
modificano, anche nell’adulto, in misura variabile da un individuo all’altro in base
all’esperienza.
Espansione della rappresentazione corticale delle dita interessate dall’addestramento.
SINDROME DELL’ARTO FANTASMA
In che misura le modificazioni delle mappe corticali influenzano la percezione? Come
vengono interpretate dal SNC?
Esperimenti di visualizzazione della corteccia somatosensitiva condotti su pazienti a cui era
stata amputata una mano, suggeriscono che le sensazioni dell’arto fantasma sono dovute
a una riorganizzazione dei circuiti corticali.
Le fibre afferenti dell’arto mancante vanno incontro a degenerazione e le vie afferenti
adiacenti si espandono e prendono il loro posto. L’aria della mano riceve afferenze da
un’altra regione cutanea: rimappatura delle sensazioni riferite.
CORTECCIA SOMATOSENSITIVA E CORTECCIA ASSOCIATIVA PARIETALE
Le aree associative parietali integrano le informazioni somatosensitive con quelle di altre
modalità sensoriali per generare sensazioni spaziali degli oggetti presenti nello spazio
extrapersonale o lontano.
Lesioni di queste aree provocano deficit complessi nella percezione dello spazio personale
o peripersonale, dell’integrazione visuomotoria e dell’attenzione selettiva.
Sindrome di negligenza personale: alterazione della consapevolezza del lato leso del
corpo.
Astereoagnosia: incapacità di riconoscere gli oggetti con il tatto.
Anosoagnosia: negazione della malattia e del deficit motorio.
SISTEMA SOMATOSENSORIALE A - B
Il sistema somatosensoriale, costituito da recettori e neuroni che si occupano di registrare e
trasmettere tutte le informazioni corporee. Si divide in sistema A e sistema B; ognuno di
essi ha una parte periferica e una centrale.
Sistema A: perifericamente comprende fibre molto mielinizzate. Le fibre si chiamano Aalfa
e Abeta.
Aα = il P.A. velocità maggiore e trasportano le informazioni propriocettive e fanno capo a 3
recettori: fusi neuromuscolari / organi tendini del golgi / recettori articolari
↳comunicano al SNC il posizionamento di alcune parti del corpo = propriocezione
Aβ = meno veloci; comunicano al cervello le sensazioni cutanee a partire da 4 recettori
cutanei.
→pacini / ruffini / messner / merkel
-questa modalità sensoriale prende nome di: esterocezione
>le fibre Aalfa e Abeta fanno sinapsi nel SNC sulle lamine 3-4 dl midollo spinale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
→dalle lamine 3-4 partono fasci gracile e cuneato che andranno a fare sinapsi nei nuclei
omonimi nel tronco encefalico.
↪ da qui le informazioni passano obbligatoriamente per il taamo, vanno prevalentemente
alla corteccia somatosensitiva primaria.
Sistema B: nella parte periferica è composto da fibre leggermente mielinizzate: Adelta e C
→ non hanno dei veri recettori, infatti terminano nel tessuto come terminazioni libere
Aδ = trasportano informazioni del danno tessutale immediato o che sta per avvenire
↳sono responsabili del 1° dolore = dolore acuto che sappiamo indicare precisamente.
[queste fibre non trasportano l’info dolorifica, esse trasportano l’info del danno
tissutale => infatti ci si riferisce a questo genere di info con il termine di nocicezione]
C= informano il S.N.C. costantemente ma in modo molto lento
→ informano il SNC di due cose: -2° dolore= dolore che insorge in ritardo e che riusciamo a
discriminare tempo dopo e non è ben definibile
-informano costantemente il cervello dello stato metabolico nelle varie parti del corpo;
informano il cervello dello stato di interocezione
>le fibre Adelta e C fanno sinapsi nelle lamine 1-2 del midollo spinale. Da queste lamine
parte il fascio spinotalamico laterale che fa sinapsi sul tronco dell’encefalo, da qui si passa
per il talamo e si arriva a livello della corteccia anteriore del cingolo e sull’insula.
PROPRIOCEZIONE
Per propriocezione (sensibilità profonda, sensibilità chinestesica) si intende la percezione
della posizione e del movimento degli arti e del corpo nello spazio.
La propriocezione possiede tre qualità:
· Senso della posizione
· Senso del movimento
· Senso della forza e della lunghezza muscolare
I propriosensori sono coinvolti in numerose attività motorie coscienti ed incoscienti.
Alla percezione della posizione del nostro corpo nello spazio partecipano tutte le
informazioni afferenti dai meccanosensori di tutti gli organi che vengono influenzati,
direttamente o indirettamente, dalle forze, dalle posizioni o dalle variazioni della posizione,
nonché dalla gravità.
PROPRIOCETTORI
Muscolo Fuso neuromuscolare
Tendine Organo muscolo-tendineo di Golgi
Articolazioni Corpuscolo di Pacini
Corpuscolo di Ruffini
Legamenti articolari Organo muscolo-tendineo di Golgi
Cute Corpuscolo di Pacini
Corpuscolo di Ruffini
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Organi vestibolari Utricolo e Sacculo
Canali semicircolari
RECETTORI MUSCOLARI E MUSCOLO-TENDINEI
I recettori localizzati all’interno dei muscoli scheletrici avvertono i cambiamenti di lunghezza
e tensione muscolare. Essi sono:
· Fusi neuromuscolari
· Organi muscolo-tendinei di Golgi
FUSO NEUROMUSCOLARE
La regione centrale o equatoriale è priva di miofibrille e non può contrarsi.
Le terminazioni dei nervi sensitivi avvolgono la regione centrale e si attivano quando la
regione equatoriale viene stirata.
Le estremità, o regioni polari, contengono miofibrille che costituiscono la parte contrattile
del fuso.
Le miofibrille si contraggono in risposta ai comandi motori dei motoneuroni gamma, che le
innervano.
Le normali fibre muscolari contrattili vengono dette anche fibre extrafusali.
Gli organi tendinei di Golgi sono posti al confine tra muscolo e tendine.
I motoneuroni gamma, il cui corpo cellulare si trova nel SNC, innervano le fibre intrafusali. I
neuroni sensoriali tonicamente attivi inviano informazioni al SNC.
Le fibre intrafusali si trovano entro i fusi neuromuscolari.
ORGANO MUSCOLO-TENDINEO DI GOLGI
È costituito da neuroni sensoriali intrecciati a fibre collagene. Se le fibre collagene vengono
stirate a seguito di accorciamento (contrazione) o allungamento (stiramento) muscolare,
esse premono/stirano le terminazioni sensitive (Ib) eccitandole e innescando i potenziali
d’azione.
Lo stimolo principale è l’accorciamento muscolare.
Le informazioni fornite da organi tendinei del Golgi e fusi neuromuscolari sono
complementari e sono utilizzate dal SNC per determinare le posizioni relative dei vari
segmenti degli arti e per la corretta esecuzione dei vari atti motori.
DOLORE
DEFINIZIONE
Il dolore è una esperienza sensoriale ed emotiva sgradevole che è collegata ad un danno
tissutale attuale o potenziale o comunque descritta nei termini di un tale danno.
Al pari di altre modalità sensitive, il dolore svolge un’importante funzione protettiva, in
quanto mette in guardia da danni ai tessuti che quindi devono essere evitati o corretti.
Tuttavia il dolore è diverso dalle altre modalità sensoriali in quanto presenta delle
caratteristiche di urgenza e primordialità che sono responsabili degli aspetti affettivi ed
emozionali della percezione del dolore.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Il carattere di spiccata individualità e soggettività del dolore costituisce uno dei fattori che
rendono difficile la sua definizione e trattamento. La percezione del dolore è soggettiva ed
influenzata da molti fattori.
INSENSIBILITÀ CONGENITA AL DOLORE
· Mancato sviluppo dei nocicettori periferici;
· Alterazione nella trasmissione sinaptica delle vie del SNC che mediano il dolore;
· Mutazioni genetiche del gene SNC9A che codifica per un unico canale voltaggio
dipendente espresso solo nei nocicettori.
Il dolore non è espressione diretta di un evento sensitivo ma il prodotto di un complesso
processo di elaborazione di segnali nervosi svolto dal SNC.
· Dolore nocicettivo fisiologico: azione di stimoli nocivi sui nocicettori di un tessuto sano.
· Dolore nocicettivo patologico: alterazione fisiopatologica di tessuti ed organi (es.
infiammazione).
· Dolore neuropatico: da attività nervosa anormale (da lesione dei nervi o del SNC,
oppure da riorganizzazione funzionale del sistema nervoso).
· Dolore nocicettivo: evocato dalla stimolazione dei nocicettori e dalla conseguente
attivazione delle fibre afferenti primarie. Es. infiammazione, stimolazione dei recettori
periferici da parte di alcuni mediatori dell’infiammazione.
· Dolore neuropatico: prodotto dal danno a diversi livelli del sistema nervoso centrale o
periferico. Es: lesione di un tronco nervoso periferico o del midollo spinale è dovuto alla
stimolazione delle vie che conducono l’informazione dolorifica.
CLASSIFICAZIONE DEL DOLORE IN BASE ALLA LOCALIZZAZIONE
Sulla base del luogo di insorgenza, il dolore può essere suddiviso in: somatico e viscerale.
· Somatico superficiale quello proveniente dalla cute es. ustione.
· Somatico profondo quello proveniente dai muscoli, dalle ossa, dalle articolazioni
e dal tessuto connettivo es. crampo muscolare
· Viscerale quello proveniente dagli organi interni es. colica renale.
I nocicettori sono rappresentati da terminazioni nervose libere localizzate nella maggior
parte del corpo.I nocicettori sono le terminazioni delle fibre afferenti primarie che portano i
segnali nocicettivi al midollo spinale. Sono state individuate tre classi principali di
nocicettori:
· Termici
· Meccanici
· Polimodali
· Nocicettori silenti
NOCICETTORI TERMICI
· Nocicettori termici: vengono attivati da livelli estremi di temperatura corporea. Sono
costituite da terminazioni periferiche di fibre mieliniche sottili Aδ che conducono i
potenziali d’azione alla velocità di 5-30m/s
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Nocicettori meccanici: vengono attivati da stimoli pressori di elevata intensità
applicati alla cute anch’essi costituiti da fibre mieliniche sottili Aδ.
· Nocicettori polimodali: possono essere attivati da stimoli meccanici, chimici, o
termici (caldo o freddo intenso). Sono costituiti da terminazioni di fibre C, di piccolo
diametro, amieliniche che conducono gli impulsi a una velocità inferiore a 1m/s.
· Nocicettori silenti: si trovano nei visceri. Non sono attivati da stimoli nocivi ma la loro
soglia viene abbassata notevolmente dai processi infiammatori e da varie sostanze
chimiche.
Le prime tre classi di nocicettori si distribuiscono estensivamente sia alla cute che ai tessuti
profondi e spesso operano insieme.
Dolore di tipo puntorio (primo dolore): immediato, acuto e ben localizzato seguito da un
senso di indolenzimento, dolore non ben definito, diffuso e sordo che a volte diventa un
vero e proprio dolore di tipo urente (secondo dolore).
Il dolore puntorio viene trasmesso dalle fibre Aδ , che ritrasmettono in modo più veloce
informazioni dai nocicettori termici e meccanici, mentre il dolore sordo, lento, viene
trasmesso dalle fibre C (segnali dai nocicettori polimodali).
Gli stimoli nocivi depolarizzano le terminazioni nervose libere afferenti e generano
potenziali d’azione che si propagano verso il SNC. Come avviene?
La membrana dei nocicettori contiene recettori che convertono l’energia termica,
meccanica e chimica degli stimoli nocivi in un potenziale elettrico depolarizzante. Una di
queste proteine fa parte di una grande famiglia proteica quella dei canali ionici del tipo TRP
(potenziale di recettore provvisorio).
· TRPM8 attivato da temperature basse e da sostanze chimiche come il mentolo.
· TRPV1 espresso selettivamente da nocicettori media le azioni dolorose della
capsaicina e di molte altre sostanze chimiche piccanti. Attivato anche da stimoli termici
nocivi e in generale è responsabile dell’insorgenza della sensazione di dolore da stimoli
termici elevati e riduzione del pH.
· TRPV2 espresso prevalentemente dalle terminazioni di Aδ ed attivato da T molto
alte.
Altri recettori e canali ionici che prendono parte alla trasduzione degli stimoli nocivi sono:
· Canali Na⁺ (Es: SCN9A una sua mutazione è responsabile dell’insensibilità
congenita al dolore;
· PTX3 (attivato da lesioni tissutali);
· recettori accoppiati a proteine G. Tutti questi diversi tipi di recettore costituiscono il
bersaglio per lo sviluppo di farmaci contro il dolore.
In numerose condizioni patologiche si possono osservare un attivazione incontrollata dei
nocicettori. Allodinia e Iperalgesia.
· Allodinia: i pazienti avvertono dolore in risposta a stimoli altrimenti innocui. Il dolore
non è costante e non è presente senza stimoli periferici.
· Iperalgesia: i pazienti hanno una risposta eccessiva agli stimoli dolorosi il dolore è
persistente anche in assenza di stimoli sensitivi.
I segnali provenienti dai nocicettori vengono trasmessi a neuroni del corno dorsale del
midollo spinale.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
L’ingresso delle fibre nocicettive avviene principalmente a livello delle lamine I,II, IV, V.
Lamina I o strato marginale risponde a stimoli nocivi che attivano direttamente fibre Aδ e C.
Rispondono selettivamente agli stimoli nocivi e vengono detti neuroni nocicettivi specifici.
Proiettano ai centri superiori e spt al talamo.
Altri neuroni secondari vengono detti ad ampio spettro dinamico e ricevono afferenze sia
dalle fibre nocicettive Aδ e C sia da Aβ, comprendendo un ampio spettro di informazioni
questi si trovano principalmente nella V lamina. I neuroni sensitivi secondari che ricevono
le afferenze non nocicettive Aβ sono invece localizzati principalmente nelle lamine II e IV.
I neuroni della lamina V ricevono afferenze anche dai nocicettori viscerali. Afferenze
nocicettive somatiche e viscerali sui neuroni della lamina V può spiegare il «dolore riferito».
DOLORE RIFERITO
Il dolore degli organi interni viene di solito percepito sulla superficie del corpo nelle regioni
indicate. Questa sensazione è definita dolore riferito.
Una teoria sull’origine del dolore riferito afferma che i nocicettori localizzati in sedi diverse
convergono su un singolo tratto ascendente del midollo spinale. Poiché i segnali dolorosi
cutanei sono più comuni di quelli derivati dagli organi interni, il cervello ha imparato ad
associare l’attivazione della via dolorifica viscerale con l’attivazione della via del dolore di
una regione cutanea. Quando la via viene attivata dal dolore in un organo interno, il
cervello associa comunque il dolore con la struttura a cui lo associa di solito, cioè la cute.
Un’altra spiegazione può essere di tipo anatomico basata sul fatto che alcuni neuroni
nocicettivi a livello periferico si ramificano e innervano sia territori cutanei sia territori
viscerali.
Il ruolo dei nocicettori non è solo quello di trasmettere informazioni afferenti ma anche
quello di secernere sostanze per la neurotrasmissione e neuromodulazione.
I nocicettori sono anche capaci di indurre una infiammazione neurogena perché dipende
dall’attività nervosa, tramite la liberazione periferica di peptidi (Sostanza P, CGRP) dalle
terminazioni periferiche delle fibre C.
SENSIBILIZZAZIONE CENTRALE
In condizione di grave danno tissutale le fibre C scaricano in modo ripetitivo e la risposta
dei neuroni delle corna dorsali aumenta progressivamente detto: caricamento e si ritiene
che per la sua genesi siano implicati recettori per il Glu (AMPA e NMDA).
Meccanismi simili a quelli che inducono potenziamento a lungo termine delle risposte
sinaptiche, che inducono cambiamenti a lungo termine della risposta dei neuroni nel corno
dorsale. Queste modificazioni prolungate dell’eccitabilità costituiscono una sorta di
memoria dello stato delle fibre afferenti C.
In alcune condizioni patologiche le alterazioni dell’eccitabilità dei neuroni del corno dorsale
possono provocare una diminuzione della soglia per il dolore e la comparsa di dolore
spontaneo → dolore dell’arto fantasma
LE VIE ASCENDENTI
Le informazioni nocicettive vengono ritrasmesse da cinque vie ascendenti:
Anterolaterale Posteriore
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Spinotalamico Cervicotalamico
↪ Laterale o Neospinotalamico
↪Mediale o Paleospinotalamico Spinoipotalamico
Spinoreticolare
Spinomesencefalico
Rivestono un ruolo minore nella nocicezione essendo deputati principalmente alla
trasmissione delle informazioni tattili. Il tratto spinoipotalamico proietta ai nuclei ipotalamici
che fungono da centri di controllo del SNA implicati nella regolazione nelle risposte
neuroendocrine e cardiovascolari che si osservano in molte sindromi dolorose.
SPINOTALAMICO
Ha un’importanza fondamentale nella trasmissione degli stimoli nocicettivi. Via più
sviluppata del midollo spinale (MS). È costituita dagli assoni di neuroni nocicettivi specifici,
termosensibili e ad ampio spettro dinamico delle lamine Ie V-VII del corno dorsale. Gli
assoni attraversano la linea mediana del midollo spinale a livello del segmento spinale da
cui originano e ascendono nel cordone anterolaterale per terminare nel talamo. Una parte
diretta al talamo mediale (paleospinotalamico) e una parte diretta al talamo laterale
(neospinotalamico).
SPINORETICOLARE E SPINOMESENCEFALICO
Spinoreticolare: assoni dei neuroni di proiezione delle lamine VII e VIII. Ascende anch’esso
nel quadrante anterolaterale e termina nella formazione reticolare e nel talamo
Spinomesencefalico: neuroni delle lamine I e V. Prende parte all’elaborazione della
componente affettiva del dolore. Decorre nel quadrante anterolaterale e proietta alla
formazione reticolare e alla sostanza grigia periacqueduttale e al nucleo parabrachiale che
proietta all’amigdala, formazione chiave per l’elaborazione degli stati emotivi.
TALAMO E NOCICEZIONE
La caratteristica principale della proiezione nocicettiva all’area S-I è l’organizzazione
somatotopica e la capacità di discriminazione dell’intensità e della durata dello stimolo. In
altre parole, questa proiezione è responsabile delle capacità discriminative, vale a dire, la
localizzazione, la durata e l’intensità di uno stimolo nocivo.
Per semplificare la complessità anatomica delle vie dolorifiche, le vie ascendenti
organizzate somatotopicamente che proiettano ai nuclei più laterali del talamo e da qui alla
corteccia S-I, vengono definite sistema laterale.
I neuroni dei nuclei mediali del talamo proiettano invece a diverse strutture, specialmente
quelle appartenenti al sistema limbico, come la corteccia orbitofrontale e il giro del cingolo,
nonché all’area somestesica (o somatosensitiva) II (S-II), all’insula e all’operculum
parietale.
Tutte queste strutture non mostrano un’organizzazione somatotopica precisa. Esse sono
responsabili della componente affettiva ed emotiva del dolore, cioè quella sensazione con
connotato negativo che fa percepire il dolore come qualcosa di spiacevole da evitare. In
altre parole, si potrebbe dire che le diverse regioni del sistema limbico, a cui i nuclei del
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
talamo proiettano, in modo particolare la corteccia cingolata anteriore, sono responsabili di
ciò che viene chiamato “sofferenza”.
Queste vie ascendenti, che passano attraverso i nuclei mediali del talamo, e che proiettano
diffusamente a diverse zone del sistema limbico, vengono definite sistema mediale.
LE AREE DEL CINGOLO E DELL’INSULA SI ATTIVANO DURANTE LA PERCEZIONE
DEL DOLORE
Il giro del cingolo fa parte del sistema limbico e si ritiene sia implicato nell’elaborazione
emozionali che accompagnano il dolore.
Asimbolia per il dolore: pazienti con lesioni dell’insula riconoscono il dolore ma non hanno
una risposta di tipo emozionale, non percepiscono nessun connotato spiacevole , tipico del
dolore. Sono in grado di tollerare stimoli dolorifici intensissimi.
Corteccia nella quale vengono elaborate le componenti sensitiva, affettiva e cognitiva del
dolore.
L’insula riceve proiezioni dirette dal talamo soprattutto da quello mediale. I neuroni
dell’insula ricevono informazioni relative ai visceri e contribuisce all’elaborazione della
componente vegetativa delle risposte agli stimoli dolorifici.
SISTEMI DISCENDENTI
Sistema di controllo discendente del dolore:
· Corteccia cerebrale
· Ipotalamo
· Tronco encefalico
La stimolazione di queste aree determina un’inibizione dei neuroni nocicettivi presenti nel
midollo spinale con diminuzione della frequenza di scarica.
Corteccia parietale e prefrontale (CCA, orbitofrontale, S-I e S-II).
Sottocorticali: amigdala, ipotalamo, talamo.
Tronco encefalico: sostanza grigia periacqueduttale, tegmento pontomesencefalico
dorsolaterale, bulbo rostroventromediale (→ rafe magno)
OPPIOIDI ENDOGENI
Le sostanze attive dell’oppio sono una classe di alcaloidi vegetali, il più abbondante dei
quali è la morfina, ancora oggi uno degli analgesici più efficaci in uso.
I ligandi endogeni per i recettori degli oppiacei, ora identificati, sono una famiglia composta
da più di venti peptidi che vengono raggruppati in tre classi:
· Endorfine
· Encefaline
· Dinorfine
In generale questi peptidi hanno una funzione depressiva.
Le endorfine sono state trovate in tutte le regioni coinvolte nella modulazione del dolore nel
bulbo rostrale ventrale ( da dove si dipartono fibre nervose discendenti eccitatorie e
inibitorie dirette ai neuroni del corno dorsale del midollo spinale) e in altre zone del midollo
spinale coinvolte nella modulazione del dolore.
Una classe di neuroni a circuito locale contenenti encefaline stabilisce sinapsi con i neuroni
che proiettano nel corno dorsale. Il rilascio di encefaline nelle terminazioni nocicettive
impedisce loro di liberare il neurotrasmettitore nei neuroni di proiezione, riducendo il livello
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
di attività che è passato ai centri superiori. Neuroni a circuito locale sono essi stessi
bersaglio delle proiezioni discendenti, provvedendo a un potente meccanismo attraverso il
quale i centri superiori possono ridurre l’attività trasmessa dalle afferenze nocicettive.
IL PRURITO
Sensazione cutanea molesta, che provoca il desiderio di grattarsi, ed evoca stimoli
meccanici, termici e chimici. La scarsa localizzabilità e la persistenza della sensazione
dopo la rimozione dello stimolo, che lo accomunano al dolore, del quale è stato per anni
considerato una submodalità.
Oggi il prurito e il dolore sono considerati due sensazioni diverse: evocano reazioni motorie
diverse, nella stessa regione cutanea il prurito è evocato da stimoli di lieve entità, il dolore
da stimoli intensi. Prurito e dolore possono essere percepiti contemporaneamente nella
stessa regione. Sembra essere mediato da una classe di fibre C, scoperta recentemente
caratterizzata da bassa velocità di conduzione.
EMOZIONI
Le emozioni sono l’insieme di risposte fisiologiche che si manifestano in modo più o meno
cosciente quando il cervello rileva situazioni particolarmente impegnative a particolari tipi di
stimoli; sono presenti in tutte le specie.
Le emozioni sono un misto di:
· attivazione fisiologica
· modificazione del comportamento
· esperienza soggettiva
Esistono emozioni di base, innate e universali:
· Presentano specifica origine funzionale
· Manifestazioni espressive specifiche
· Modificazioni fisiologiche
· Prevedibili tendenze all’azione
LE EMOZIONI SONO UN PRODOTTO EVOLUZIONISTICO
· Valutazione degli eventi ambientali
· Rivalutazione di scopi e obiettivi
· Preparazione all’azione
La muscolatura facciale è controllata:
· Movimento volontario corteccia motoria primaria
· Sistema nervoso delle emozioni corteccia del cingolo e prefrontale
LE ESPRESSIONI FACCIALI: GLI ESPERIMENTI DI DUCHENNE
Usando uno stimolatore elettrico, applicato a determinati punti della pelle, riuscì a
catalogare tutti i muscoli della faccia umana. Si interessò anche al modo in cui i muscoli
facciali vengono utilizzati per trasmettere emozioni. A seconda che siamo arrabbiati, felici,
annoiati, spaventati, il nostro volto esprime questi stati d’animo con un complesso insieme
di movimenti muscolari; Duchenne riuscì a “comporre” e catalogare queste espressioni
emozionali.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
SORRIDERE CON GLI OCCHI NON È SOLO UNA METAFORA
Nel “sorriso di cortesia”, i muscoli attorno alla bocca si attivano in maniera molto simile al
sorriso naturale. Questo sorriso “bugiardo” difetta però di qualcosa: sono i muscoli intorno
agli occhi e alle sopracciglia, scoprì Duchenne, che si muovono soltanto se il sorriso è
sincero e genuino.
L'encefalo elabora le espressioni facciali.
MODELLO DI JAMES-LANGE
L'esperienza di una emozione è la consapevolezza delle risposte fisiologiche agli stimoli
capaci di suscitarla. Si propone che l’esperienza emotiva cosciente derivi dai cambiamenti
fisiologici nel corpo. IL corpo crea l’emozione → l’info dal corpo raggiunge la corteccia →
nella corteccia si crea l’emozione cosciente
>l’esperienza emotiva cosciente avviene dopo che la corteccia ha ricevuto informazioni
circa le modificazioni fisiologiche.
CANNON - CRITICHE A JAMES
Emozioni diverse correlano con reazioni fisiologiche identiche; gli stati fisiologici non
possono differenziare le emozioni. I cambiamenti fisiologici sono generalmente troppo lento
per generare un’emozione e l’attivazione fisiologica tramite ormoni non induce emozioni.
MODELLO DI CANNON-BARD
Gli stimoli capaci di suscitare emozione innescano allo stesso tempo:
· risposte fisiologiche
· esperienza personale della emozione
Per lui l’esperienza emotiva e l’espressione emotiva non sono due variabili che si
influenzano - ma sono due entità separate.
le info sensoriali giungono → passa all'ipotalamo che darà
al talamo vita ai comandi motori che => reazione fisiologica
↓ producono le emozioni collegata all’emozione
proietta anche alla corteccia
cerebral che darà origine
all’emozione cosciente
IPOTALAMO
Ruolo strategico e di particolare rilievo sul controllo delle funzioni vegetative. Esso riceve
segnali dal tutto il corpo che riguardano informazioni: viscerali, visive, termiche, dolorifiche.
Dall’interno delle strutture ipotalamiche nascono inoltre segnali relativi alla temperatura del
sangue, all’osmolarità plasmatica, alla concentrazione ematica di glucosio e di molti
ormoni. L’insieme delle informazioni interne e/o afferenti verrebbero poi confrontati con i
diversi livelli di riferimento per far scaturire risposte intese a mantenere l’omeostasi di
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
parametri fisiologici quali: temperatura corporea, pressione sanguigna, equilibrio idrosalino
e metabolismo energetico.
Stimolazione di nuclei diversi dell’ipotalamo inducono sensazioni di:
· rabbia
· paura
· piacere
Nucleo laterale: controlla le risposte viscerali e somatiche del comportamento emotivo
Nucleo paraventricolare: controlla le risposte ormonali
INTEGRAZIONE DELLE INFORMAZIONI: L’IPOTALAMO
Informazioni contestuali
· Corteccia cerebrale
· Amigdala
Informazioni sensoriali
· Vie sensoriali somatiche e viscerali
· Segnali umorali e chemiocettivi
↓
Ipotalamo: confronta gli input con i valori di omeostasi
↓
Risposte motorie viscerali, motorie somatiche, neuroendocrine e comportamentali
STRUTTURE EFFETTRICI
Sistema Nervoso Autonomo o Vegetativo è quella parte del sistema nervoso che provvede
alla regolazione delle funzioni viscerali dell’organismo.
Esso è essenzialmente un sistema effettore che presiede al controllo della muscolatura
liscia, di quella cardiaca e delle funzioni ghiandolari. Coadiuvato in molti casi dal sistema
endocrino, esplica una funzione modulatrice sulla gittata cardiaca e sulla pressione
arteriosa, sull’attività gastrica e intestinale, sulle secrezioni ghiandolari e su molte altre
funzioni viscerali, ha un elevata velocità di reazione, essendo capace di rispondere nel giro
di pochi secondi alle variazioni dell’ambiente interno, garantendo un rapido ripristino delle
condizioni di stabilità.
↓
Omeostasi:
Tendenza naturale al raggiungimento di una relativa stabilità (proprietà chimico-fisiche
interne che comportamentali), che accomuna tutti gli organismi viventi, per i quali tale
regime dinamico deve mantenersi nel tempo, anche al variare delle condizioni esterne,
attraverso precisi meccanismi autoregolatori.
OMEOSTASI
Il mantenimento della costanza della composizione dell’ambiente interno dell’organismo -
omeostasi - è una delle principali funzioni del sistema nervoso autonomo (SNA).
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
La regolazione omeostatica è mediata dal SNA in congiunzione con l’ipotalamo ed altri
centri a livello cerebrale e si basa su un meccanismo che mantiene una variabile a valori
costanti o entro precisi limiti di variazione.
SISTEMA NERVOSO AUTONOMO
Il Sistema Nervoso Autonomo (SNA), è la parte del sistema nervoso periferico motorio e
sensitivo che regola quelle attività corporee che generalmente non sono sotto il controllo
della coscienza. Il SNA è involontario.
Il sistema nervoso autonomo innerva tutti gli organi, producendo prevalentemente azioni
involontarie e automatiche che sono mediate da due sezioni principali, il simpatico e
parasimpatico, che sono distinte da un punto di vista neurochimico e anatomico. Numerosi
connessioni sinaptiche e neurotrasmettitori consentono il controllo autonomo della funzione
dell'organo a livello locale e centrale in maniera integrata, per venire incontro alle esigenze
di tutto il corpo.
SUDDIVISIONE DEL SNA
· Sistema simpatico (ortosimpatico)
· Sistema parasimpatico
· Sistema enterico (metasimpatico)
Il sistema simpatico si attiva in contesti di allerta, emergenza, emozione, attività fisica,
stress…una sua attivazione generalizzata media la reazione istintiva ad un pericolo detta di
attaco o fuga (fight-or-flight reaction).
Il parasimpatico è maggiormente attivo nelle condizioni di riposo e durante le fasi digestive:
un sistema responsabile del rest and digest (Cannon).
SI POSSONO MISURARE LE EMOZIONI?
Macchina della verità
· respirazione toracica e addominale
· sudorazione
· attività cardiovascolare
CIRCUITO DI PAPEZ
le aree che fanno parte di questo circuito sono la:
corteccia del cingolo / ippocampo / ipotalamo/ nuclei talamici anteriori
↪sede dell’esperienza emotiva ↪sede dell’espressione emotiva
>realizza una connessione bidirezionale tra la corteccia cingolata e l’ipotalamo
INSULA
Coinvolta negli aspetti valutativi espressivi degli strati emotivi generati internamente.
Lesioni alla corteccia dell’insula, soprattutto emisfero sinistro, possono comportare la
scomparsa temporanea di comportamenti dovuti a stati di dipendenza. Il ruolo funzionale
nei processi di associazione fra segnali esterni ed interni come il desiderio e il piacere.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
AMIGDALA
Organizza le risposte comportamentali vegetative e ormonali di rabbia paura ed ansia. È
implicata nei comportamenti sessuali e materni.
È costituita da nuclei basolaterali che ricevono informazioni da corteccia e talamo, da un
nucleo mediale che riceve input olfattivi dal bulbo olfattivo ed accessorio, e da un nucleo
centrale che invia comandi alle strutture motorie del tronco encefalico ed all'ipotalamo
(funzioni vegetative e comportamenti specie specifici).
APPRENDIMENTO E PAURA CONDIZIONATA
· Comportamento di immobilizzazione (PAG)
· Risposte del SNA (ipotalamo laterale)
· Secrezione degli ormoni dello stress (nucleo paraventricolare ipot. sistema ipot.,
ipofisi-corticale surrene)
Lesioni dei nuclei basolaterali dell'amigdala aboliscono la capacità di associare il tono con
la scarica elettrica. L'animale non si immobilizza più né mostra alterazioni fisiologiche in
risposta al tono, sebbene continui a mostrare una normale risposta fisiologica alla scarica
elettrica.
LA PAZIENTE S.M.- MALATTIA DI URBACH-WIETHE
Calcificazione bilaterale dell’amigdala. La paziente non è in grado di riconoscere lo stato
emotivo corrispondente alla paura.
LA SINDROME DI KLÜVER–BUCY
Lesione bilaterale del lobo temporale mediale (inclusi i nuclei amigdaloidei):
· Iperfagia
· Ipersessualità
· Iperoralità
· Agnosia visuale
DISTINGUIAMO LE EMOZIONI DAI SENTIMENTI
Sentimenti: esperienza cosciente delle emozioni. Sorta di resoconto che il cervello crea per
rappresentare i fenomeni fisiologici generati dallo stato emozionale.
I sentimenti complessi sono associati con interazioni sociali che comprendono una serie di
sentimenti (empatia, orgoglio, imbarazzo, senso di colpa etc.).
Importanti contributi alle normali interazioni sociali.
CORTECCIA CINGOLATA ANTERIORE
· La ACC è coinvolta nella regolazione di funzioni autonomiche ed endocrine,
valutazione della salienza di informazioni emozionali e nella regolazione delle risposte
emotive.
· Coinvolta nel comportamento motivato grazie al circuito che la mette in connessione
con i gangli della base (circuito limbico).
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· La ACC è coinvolta nell’attività di mediazione dell’attivazione emozionale, soprattutto
quando l’informazione esterna è in contrasto con lo stato interno.
· La ACC è particolarmente attiva durante compiti di richiamo indotto di un emozione,
ad esempio attraverso la rievocazione di memorie o immagini di eventi autobiografici
rilevanti.
CORTECCIA PREFRONTALE
Pazienti con questo tipo di lesioni non riescono a mantenere il posto di lavoro, non riescono
a intrattenere relazioni sociali stabili, violano le convenzioni sociali e non riescono a
mantenersi indipendenti dal punto di vista finanziario.
In condizioni sperimentali controllate, le valutazioni morali di questi pazienti possono
essere incrinate.
IL CASO DI PHINEAS GAGE:
Gage sopravvisse 12 anni, ma con gravi alterazioni della personalità.
“Egli è sregolato, irriverente, indulge talvolta nella bestemmia più volgare (che in
precedenza non era suo costume), manifestando poco rispetto per i suoi compagni,
intollerante verso limitazioni o avvertimenti quando questi vanno in conflitto con i suoi
desideri, talora tenacemente ostinato, capriccioso ed esitante, progetta molti piani per il
futuro, che vengono tuttavia abbandonati, anziché essere organizzati, in favore di altri piani
che sembrano più facilmente attuabili […].”
1936: INIZIA L’ERA DELLA PSICOCHIRURGIA
Usi della lobotomia frontale
LATERALIZZAZIONE CORTICALE DELLE FUNZIONI EMOTIVE
Emisfero destro maggiormente coinvolto nell’espressione e comprensione dei connotati
emotivi:
· del linguaggio (comprensione emotiva attraverso il linguaggio)
· delle espressioni facciali
Emisfero sinistro: emozioni positive
Lesioni emisfero anteriore sinistro:
· Perdita di emozioni positive
· Grave depressione
Emisfero destro: emozioni negative
Lesioni emisfero anteriore destro:
· Perdita di emozioni negative
· Eccessivo ottimismo
TURBE DEGLI UMORI E GLI STATI ANSIOSI
Quando uno stato emotivo si prolunga, può divenire, col tempo, lo stato emotivo
dominante: il nostro umore. L’umore può essere indipendente dalle circostanze immediate
sia personali che ambientali.
Depressione:
· Umore depresso
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Perdita di interesse o di piacere
· Perdita/aumento di peso
Stati ansiosi:
· Ansia generalizzata (diffusa, subcontinua, cronica, non legata a stimoli specifici)
· Attacchi di panico (acuta, attacchi, dovuta a stimoli specifici)
LINGUAGGIO
Il linguaggio è una caratteristica esclusiva dell’uomo ed è la nostra capacità più elevata e la
nostra massima acquisizione.
Il linguaggio verbale rappresenta la facoltà mentale che ci permette di codificare le proprie
idee e stati d’animo in simboli intelligibili, come suoni articolati, segni grafici e gesti, a scopo
di comunicazione interpersonale → vita sociale.
Sembra che il linguaggio verbale sia una funzione mentale che interessi la sola specie
umana; negli animali inferiori esistono pure forme di comunicazione sociale ma esse
apparentemente mancano di un contenuto simbolico astratto e dell’arbitrarietà,
caratteristiche tipiche del linguaggio umano.
ALTRE FORME DI COMUNICAZIONE
· La danza delle api per segnalare la distanza, la direzione e la quantità di cibo.
· Marcare il territorio;
· Attirare e conquistare le femmine della stessa specie;
· Avvisare i membri della stessa specie e i piccoli di minacce e pericoli;
· Mantenere i contatti con lo stormo o altri individui.
LINGUAGGIO
Una delle più straordinarie funzioni corticali nell'uomo è la capacità di associare simboli
arbitrari a significati specifici per esprimere pensieri ed emozioni a noi stessi e agli altri
attraverso il linguaggio scritto e parlato. L’eloquio e la scrittura sono solo due dei possibili
sistemi di codificazione linguistica.
APPARATO VOCALE
Gli organi che producono la parola includono:
· i polmoni, che fungono da riserva d'aria;
· la laringe, che è la sede della qualità degli stimoli periodici che costituiscono i
suoni delle parole;
· la faringe, le cavità orali e nasali e le loro strutture incluse (lingua, denti e
labbra), che modificano (o filtrano) i suoni emessi dall’oratore.
la laringe è la "fonte" dei suoni vocali e il resto del tratto vocale funge da filtro che modula
l'energia sonora della sorgente.
Fonazione: emissione della voce
Fonetica: articolazione dei fonemi
FONAZIONE
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Energia dell’aria emessa da un’espirazione fa vibrare le corde vocali, tese tra la cartilagine
tiroide e delle cartilagini aritenoidi della laringe e intercettano tra di loro una fessura: la
glottide.
L’aria passa attraverso la glottide, durante la respirazione è tenuta aperta dalla contrazione
dei muscoli cricoaritenoidei posteriori, durante la fonazione vi è la contrazione dei muscoli
cricoaritenoidei laterali (restringimento) e aritenoidei (chiusura).
La contrazione dei muscoli intrinseci della laringe dipende dal nervo laringeo inferiore,
ramo terminale del nervo ricorrente del vago, per i muscoli cricotiroidei sono innervati dal
nervo laringeo superiore (ugualmente un ramo del vago).
FONETICA
La fonetica è produzione e percezione di suoni linguistici (fonemi), e le loro caratteristiche.
L’articolazione dei fonemi avviene durante il transito dell’aria emessa dalla glottide
nell’apparato di risonanza formato da faringe, bocca e naso (tratto vocale), che
costituiscono cavità di ampiezza variabile in relazione alla posizione della lingua.
I fonemi vengono prodotti perché porzioni diverse del tratto vocale entrano in risonanza con
alcune frequenze emesse dalla glottide, dette formanti, che vengono rafforzate in intensità.
Le diverse porzioni del tratto vocale cambiano di configurazione con la posizione della
mandibola, lingua denti labbra, a ogni configurazione corrispondono diverse frequenze
formanti. Formazioni di consonanti (gutturali, labiali, dentali).
· Faringe, bocca, naso, denti, lingua
· I muscoli del tratto vocale sono innervati dai nervi trigemino, facciale,
glossofaringeo, vago, accessorio e ipoglosso.
AREE DEL LINGUAGGIO
Area di Broca / Area di Wernicke → presenti nell'emisfero sinistro
↓ ↓
44-45 22
risiede nel lobo frontale / risiede nel lobo temporale
A.di Broca: connessa al programma motorio del linguaggio
A. di Wernicke: connessa alla percezione del linguaggio
-sono connesse tra di loro attraverso un fascicolo: fascicolo arcuato
IL PAZIENTE LEBORGNE
molteplici problemi neurologici, presentava gravi problemi al linguaggio che durarono fino
alla sua morte. Ogni volta che tentava di pronunciare una frase o rispondere a una
domanda, poteva solo produrre un'unica sillaba ripetitiva, "Tan", che ripeteva di solito due
volte di seguito L'autopsia rivelò una lesione sulla superficie del lobo frontale sinistro, come
sospettava Broca.
IL PAZIENTE LELONG
In seguito ad un ictus di mostrava difficoltà nel discorso produttivo. Questo paziente di 84
anni poteva dire solo cinque parole. L'autopsia rivela anche una lesione
approssimativamente nella stessa regione del lobo frontale e Broca conferma la
localizzazione del linguaggio in quest'area. Broca capì che l'integrità della terza
convoluzione frontale sembrava indispensabile per l'esercizio della facoltà del linguaggio
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
articolato. La lesione del paziente occupava esattamente la stessa sede immediatamente
dietro la parte mediale, di fronte all'insula.
IL PAZIENTE LEBORGNE: UN’ANALISI MODERNA
Una parte della lesione era chiaramente visibile nel giro frontale inferiore, in particolare nel
terzo medio con un'ulteriore atrofia nel terzo posteriore, appena sopra la fessura silviana.
Inoltre, è stata osservata una modesta atrofia nel giro frontale medio e nel giro temporale
superiore. A controllo ravvicinato, sono stati osservati anche giri deformati e necrotici nel
lobo parietale anteriore inferiore. L'emisfero destro, era intatto. Questi risultati sono in gran
parte coerenti con la descrizione originale di Broca della lesione all'autopsia.
Le immagini mostrano un danno significativo in tutto l'emisfero sinistro, sia corticale che
subcorticale. L'emisfero sinistro è chiaramente più piccolo e distorto a causa della
distruzione della corteccia e della materia bianca in tutto l'emisfero. Le sezioni sagittale,
assiale e coronale attraverso il cervello rivelano lesioni nel giro frontale inferiore sinistro,
lobo parietale inferiore profondo e lobo temporale anteriore.
Inoltre, vi è un ampio coinvolgimento sottocorticale compreso il claustrum, il putamen, il
globus pallidus, la testa del nucleo caudato. L'intera lunghezza del fascicolo longitudinale
superiore è danneggiata. L'emisfero destro non è danneggiato e serve come un eccellente
confronto con l'emisfero sinistro.
IL PAZIENTE LELONG: UN’ANALISI MODERNA
La corteccia è gravemente atrofizzata. La fessura silviana è così allargata che l'insula è,
anormalmente, quasi completamente esposta. Le sezioni sagittale (S), assiale (A) e
coronale (C) confermano la grave atrofia e la lesione nella parte posteriore dell'area di
Broca sul giro frontale inferiore. La lesione coinvolge la parte posteriore della pars
opercularis, mentre la metà anteriore di questa struttura e l'intera pars triangularis sono
completamente risparmiate.
Inoltre, lesioni piccole ma distinte sono presenti nel fascicolo longitudinale superiore sopra
l'insula e lateralmente al corno anteriore del ventricolo laterale sinistro. L'insula, benché
gravemente atrofizzata, non è specificamente lesionata, né altre strutture profonde.
OLD BRAINS, NEW IDEAS
La relazione tra l'afasia di Broca e l'area di Broca continuò a essere studiata da numerosi
ricercatori e clinici. Il tipo di disturbo descritto da Broca non era sempre correlato alle
lesioni nella zona di Broca, ma piuttosto a lesioni che coinvolgono più strutture mediali.
Studi sulle lesioni moderne hanno scoperto che l'afasia di Broca è causata da grandi lesioni
che comprendono non necessariamente l'area di Broca, ma circondano la corteccia
frontale, la sostanza bianca sottostante, l'insula, i gangli della base e le parti del giro
temporale superiore anteriore oltre al fascicolo longitudinale arcuato / superiore. Ciò
implica che anche altre regioni del cervello partecipano alla produzione del linguaggio oltre
all'area di Broca.
ANALISI DISTRIBUITA
Wernicke ipotizzò che il programma motorio, che coordina i movimenti della bocca
necessari per l’emissione corretta della parole, sia localizzato nell’ area di Broca, che è
particolarmente ben situata per questa funzione in quanto è posta immediatamente davanti
all’area motoria che controlla la bocca, lingua, palato e corde vocali.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Localizzò invece il programma sensitivo, che controlla la percezione delle parole, nell’area
del lobo temporale da lui scoperta. Anche quest’area è localizzata in modo opportuno, in
quanto circondata dalla corteccia uditiva e da altre aree associative che integrano
informazioni acustiche visive e somatiche.
MODELLO DELL’ORGANIZZAZIONE DEL LINGUAGGIO
Le informazioni acustiche e visive che riguardano il linguaggio si formano in aree sensoriali
diverse. Da queste aree raggiungono il giro angolare che costituisce una area corticale
associativa specializzata per le informazioni di natura visiva e acustica nella quale le parole
dette o scritte vengono trasformate in un codice nervoso unico e comune sia al linguaggio
che alla scrittura. Così codificata questa informazione viene trasferita nell’area di Wernicke
e associata al proprio significato. Da qui la rappresentazione neurale viene trasferita
nell’area di Broca che custodisce le regole grammaticali necessarie per trasformare la
rappresentazione sensitiva in rappresentazione motoria che può estrinsecarsi come
linguaggio scritto o parlato.
AFASIA DI CONDUZIONE
Essa si origina a causa di lesioni delle fibre che connettono l’area di Wernicke con l’area di
Broca (fascicolo arcuato). Questi pazienti sono in grado di capire le parole che sentono e
non hanno difficoltà motorie nell’emissione di parole, ma non riescono a parlare
correttamente.
Caratteristiche dell’afasia di conduzione:
· eloquio fluente
· comprensione discreta
· difficoltà a ripetere frasi (dovuta alla disconnessione tra il centro della
produzione e quello della comprensione del linguaggio)
Nuove evidenze:
· Lesioni del giro temporale superiore sx (AB 39)
· Lobo parietale inferiore (AB 40)
· Area Spt localizzata tra area 39 e 40 fornisce sia risposte motorie che uditive
NORMAN GESCHWIND
Uno sforzo per perfezionare la categorizzazione delle afasia del diciannovesimo secolo fu
intrapreso dal neurologo americano Norman Geschwind negli anni '50 e nei primi anni '60.
Basandosi su dati clinici e anatomici provenienti da un gran numero di pazienti e su una
migliore comprensione della connettività corticale ricavata da studi sugli animali,
Geschwind ha concluso correttamente che molte altre regioni delle cortecce parietale,
temporale e frontale sono criticamente coinvolte nel linguaggio.
Fondamentalmente, ha dimostrato che il danno a queste aree aggiuntive porta a deficit
linguistici identificabili, anche se più sottili.
Il suo chiarimento delle definizioni dei disturbi del linguaggio è stato in gran parte
confermato dall'imaging cerebrale funzionale in soggetti normali e rimane la base per molti
lavori clinici contemporanei su linguaggio e afasia.
AFASIE
Afasia di Broca (motoria, espressiva o di produzione):
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
= il paziente avrà difficoltà nell’eloquio, non presenta difficoltà nella comprensione
del linguaggio.
Afasia di Wernicke (afasia sensoriale o ricettiva):
= il paziente avrà un eloquio fluente, scambierà sillaba e parole tra loro. presenta un
deficit nella comprensione del linguaggio.
Afasia motoria transcorticale:
Lesioni che interessano l’area frontale dorsolaterale di sinistra lembo di corteccia
associativa situata anteriormente e superiormente all’area di Broca (anche se la stessa
area di Broca può essere danneggiata). Importante per indirizzare l’attenzione e per il
mantenimento di talune facoltà esecutive di ordine superiore tra cui la scelta della parola
migliore.
Area motoria supplementare di sinistra, localizzata nella parte alta del lobo frontale
immediatamente al davanti dell’area motoria primaria. È attiva nei compiti che richiedono
l’emissione di parole.
· SMA contribuisce all’inizio del discorso
· Fronto-dorso-laterale contribuisce al controllo del discorso in atto
Afasia transcorticale sensoriale:
Pazienti parlano in modo fluente, ma hanno una ridotta comprensione del significato del
discorso, difficoltà nel denominare gli oggetti . La ripetizione delle parole è conservata,
possono anche suggerire correzioni grammaticali anche se non capiscono il significato.
Lesioni circoscritte dell’area di giunzione fra i lobi temporali parietale e occipitale che
connette le aree perisilviane del linguaggio con le regioni corticali che presiedono alla
comprensione del significato delle parole.
Afasia globale:
Perdita di ogni capacità di comprensione, ripetizione e fluenza. I discorsi sono ridotti all’
emissione di qualche parola. I pazienti ripetono la stessa parola, sia essa appropriata o
meno nel vano tentativo di comunicare ciò che pensano. Possono ancora pronunciare in
modo automatico espressioni di uso corrente, sono in grado di contare o elencare i giorni
della settimana.
La comprensione della parola udita si limita a poche parole ed espressioni comuni. Si
accompagna a paresi della parte destra della faccia ed è dovuta a:
· Lesioni parte anteriore del linguaggio
· Nuclei della base e dell’insula
· Lesioni delle aree corticali uditive
· Lesioni nelle regioni posteriori del linguaggio
ZONE ATTIVATE IN DIVERSI COMPITI LINGUISTICI
Le attività linguistiche come ascoltare le parole e generare associazioni di parole suscitano
l'attività nelle aree di Broca e Wernicke, come previsto.
Attività nelle aree sensoriali e motorie primarie e di associazione per compiti linguistici attivi
e passivi. Queste osservazioni indicano che l'elaborazione del linguaggio coinvolge regioni
corticali oltre alle aree linguistiche classiche.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
IL LINGUAGGIO È SIA LOCALIZZATO SIA LATERALIZZATO
La rappresentazione ineguale delle funzioni linguistiche nei due emisferi cerebrali fornisce
un esempio particolarmente convincente del fenomeno di localizzazione e lateralizzazione
di una funzione cognitiva.
La sua lateralizzazione ha dato origine all'idea fuorviante che un emisfero nell'uomo sia
effettivamente "dominante" sull'altro, cioè l'emisfero in cui risiede la maggiore capacità di
linguaggio.
Il vero significato della lateralizzazione per il linguaggio o qualsiasi altra capacità cognitiva,
tuttavia, risiede nella suddivisione efficiente di funzioni complesse tra gli emisferi, piuttosto
che in qualsiasi superiorità di un emisfero rispetto all'altro.
LATERALIZZAZIONE DEL LINGUAGGIO
Funzioni linguistiche primarie si trovano nell'emisfero sinistro:
· i collegamenti tra i suoni della parola e i loro significati sono rappresentati
principalmente nella corteccia temporale sinistra;
· la circuiteria per i comandi motori che organizzano la produzione di un
linguaggio significativo si trova principalmente nella corteccia frontale sinistra.
Nonostante questo predominio sinistro per gli aspetti "lessicali" del linguaggio, il contenuto
emotivo (affettivo) del discorso è governato in gran parte dall'emisfero destro.
Lesioni all’emisfero destro corrispondenti all’area di Broca e di Wernicke determinano
aprosodie: perdita del ritmo, accentuazione e variazioni di tono del discorso. Incapacità di
esprimere la ricchezza di sfumature del linguaggio usato ogni giorno.
Stimoli visivi o semplici istruzioni possono essere impartite indipendentemente all'emisfero
destro o sinistro in soggetti normali e con cervello diviso. Poiché il campo visivo sinistro è
percepito dall'emisfero destro (e viceversa), un'istruzione brevemente presentata
(tachistoscopica) nel campo visivo sinistro è apprezzata solo dal cervello destro. Nei
soggetti normali, l'attivazione della corteccia visiva destra porta al trasferimento emisferico
delle informazioni visive attraverso il corpo calloso all'emisfero sinistro. Nei pazienti con
cervello diviso, le informazioni presentate al campo visivo sinistro non possono raggiungere
l'emisfero sinistro e i pazienti non sono in grado di produrre un rapporto verbale riguardante
gli stimoli. Tuttavia, tali pazienti sono in grado di fornire una relazione verbale degli stimoli
presentati nel campo visivo corretto.
Conferma della specializzazione emisferica per la lingua ottenuta studiando individui in cui
le connessioni tra emisfero destro e sinistro sono state divise chirurgicamente.
Lo stereognosia (riconoscimento tattile) con una sola mano di oggetti celati alla vista può
essere utilizzata per valutare le capacità linguistiche di ciascun emisfero nei pazienti con
cervello diviso. Gli oggetti tenuti nella mano destra, che forniscono informazioni sensoriali
somatiche all'emisfero sinistro, sono facilmente nominati; gli oggetti tenuti nella mano
sinistra, tuttavia, non sono facilmente nominati da questi pazienti.
Utilizzando l'emisfero destro, i soggetti potevano produrre solo una descrizione indiretta
dell'oggetto che si basava su parole e frasi rudimentali piuttosto che sul preciso simbolo
lessicale dell'oggetto (ad esempio, "una cosa rotonda" invece di "una palla"), e alcuni non
sono stati in grado di fornire alcun resoconto verbale di ciò che hanno tenuto nella mano
sinistra.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
FUNZIONI DELL’EMISFERO DESTRO E DELL’EMISFERO SINISTRO
Una vasta gamma di funzioni emisferiche può essere valutata usando il metodo
tachistoscopico (stimolo presentato brevemente nel campo visivo), anche in soggetti
normali.
Test di Wada un anestetico a breve durata d'azione viene iniettato nella carotide sinistra;
questa procedura transitoriamente "anestetizza" l'emisfero sinistro e quindi mette alla prova
le capacità funzionali della metà interessata del cervello.
Se l'emisfero sinistro è effettivamente "dominante" per il linguaggio, allora il paziente
diventa transitoriamente afasico mentre svolge un compito verbale continuo come il
conteggio.
DIFFERENZE ANATOMICHE TRA DESTRA E SINISTRA
Le differenze nella funzione linguistica tra gli emisferi sinistro e destro hanno naturalmente
ispirato neurologi e neuropsicologi a trovare un correlato strutturale di questa
lateralizzazione comportamentale.
Una differenza emisferica che ha ricevuto molta attenzione nel corso degli anni è stata
identificata alla fine degli anni '60 da Norman Geschwind e dai suoi colleghi. Hanno trovato
un'asimmetria nell'aspetto superiore del lobo temporale noto come planum temporale.
Questa area era significativamente più grande sul lato sinistro in circa due terzi dei soggetti
umani studiati dopo la morte.
ESTENSIONE DEL PLANUM TEMPORALE NELL’EMISFERO SINISTRO
Poiché il planum temporale è vicino alle regioni del lobo temporale che contengono aree
corticali essenziali per il linguaggio, inizialmente è stato suggerito che questa asimmetria
verso sinistra riflettesse il maggiore coinvolgimento dell'emisfero sinistro nel linguaggio.
MEMORIA
DEFINIZIONI
è una funzione del SNC, non ha una precisa sede all'interno dell'encefalo, guida il
comportamento a interagire con l'ambiente.
è un meccanismo di codifica, immagazzinamento e recupero delle informazioni apprese.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Apprendimento: processo attraverso il quale il sistema nervoso acquisisce
nuove informazioni, utilizzate poi per produrre cambiamenti del comportamento.
APPRENDIMENTO
Un cambiamento del comportamento relativamente permanente che si verifica in risposta
all’esperienza.
Apprendimento non associativo (= 1 solo stimolo):
· Abituazione (o assuefazione)
· Sensibilizzazione
Apprendimento associativo (= relazione tra eventi):
· Condizionamento Classico (o associativo propriamente detto) è basato
sull’associazione di stimoli
· Condizionamento Operante (o strumentale) è basato sull’associazione
stimolo-risposta-conseguenze
DIVERSE FORME DI APPRENDIMENTO
· Abitudine: processo che provoca nell’animale una diminuzione delle risposte
indotte da una stimolazione ripetuta. Riflette la capacità del sistema nervoso di
cambiare la sua risposta agli stimoli ambientali (esempio: rumore ripetuto,
risposta tattile dei vestiti sulla pelle)
· Sensibilizzazione: aumento dell’ampiezza della risposta a uno stimolo dopo aver
ricevuto uno stimolo di tipo diverso.
CONDIZIONAMENTO CLASSICO
Capacità di produrre una risposta nuova a uno stimolo precedentemente neutro (stimolo
condizionato, SC) dopo che SC è stato abbinato ripetutamente a uno stimolo che induce
sempre la stessa risposta (stimolo incondizionato, SI).
Formazione di una relazione predittiva fra due stimoli piuttosto che la loro semplice
sequenza.
BLINK REFLEX CONDIZIONATO
Lesione del verme cerebellare abolisce la risposta condizionata (es. ammiccamento in
risposta ad un suono) ma non ha effetto sulla risposta non condizionata (ammiccamento in
risposta ad un soffio d’aria). Aumento dell’ attività dei neuroni durante lo sviluppo del
comportamento condizionato.
Lesione del nucleo interposito abolisce anche il riflesso condizionato di ammiccamento.
↓
Condizionamento classico dell’ammiccamento e forse in altre forme semplici di
condizionamento classico che comportano movimenti dei muscoli scheletrici
CONDIZIONAMENTO OPERANTE
Il soggetto deve operare sull’ambiente per ottenere un risultato.
Formazione di una relazione predittiva fra un’azione e un certo comportamento.
Si basa principalmente sul concetto di rinforzo positivo il quale andrà inevitabilmente ad
incidere sul comportamento dell’individuo.
DIVERSI TIPI DI MEMORIA
Memoria sensoriale → Memoria di lavoro (o memoria breve) ↔ Memoria a lungo termine
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Memoria sensoriale: gestisce un immenso numero di afferenze è inconscia e possiede
una grande e possiede una grande capacità è di bravissima durata.
· Memoria iconica (informazioni visive)
· Memoria ecoica (informazioni acustiche)
· Memoria aptica (informazioni tattili)
↓ focus
Memoria di lavoro (o memoria breve): ha una capacità limitata ed è l’unica che si
manifesta nell’immediato ad un livello consapevole.
· Esecutivo centrale
· Loop fonologico
· Buffer episodico
· Sketchpad visuo-spaziale
↓storage ↑retrivial
Memoria a lungo termine: inconscia, capacità enorme, ma non illimitata sua volta si
suddivide in dichiarativa e non dichiarativa
· Esplicita (dichiarativa)
o Episodica
o Semantica
· Implicita (non-dichiarativa)
o Procedurale
La memoria a lungo termine mantiene solo le informazioni ben apprese e praticate
Es.
· so andare in bicicletta → memoria motoria
· andare in bicicletta dopo 10 anni → memoria procedurale
· ho paura della bicicletta ( non so perché) → memoria emotiva
· cos’è una bicicletta → memoria semantica
· ieri sono andato in bicicletta → memoria episodica
MEMORIA DICHIARATIVA
· Memoria semantica → Memoria degli avvenimenti
· Memoria episodica → Memoria autobiografica
Lobo temporale mediale
Codifica: compiuta prestando molta attenzione alle informazioni che arrivano e
associandole a quelle già presenti
Conservazione: meccanismi neurali e siti nei quali la memoria viene conservata nel tempo
Consolidamento: rende persistenti le informazioni conservate temporaneamente
Richiamo: processo attraverso il quale le informazioni conservate vengono ricordate
IL PAZIENTE H.M.
Nel 1957 H. M. fu trattato chirurgicamente per epilessia intrattabile del lobo temporale con
asportazione bilaterale dell’ippocampo e dell’amigdala e parte della corteccia temporale.
H.M. soffrì da quel momento di un grave disturbo della memoria a breve termine. Memoria
a lungo termine per eventi avvenuti prime dell’intervento erano intatti.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Memoria semantica intatta (manteneva un buon linguaggio e ricchezza di vocaboli).
Memoria procedurale intatta (compito della stella allo specchio).
Dopo molti anni dall’intervento il paziente fu sottoposto a RM encefalo.
Il suo caso dimostrò in modo conclusivo il ruolo fondamentale della parte mediale del lobo
temporale nella memoria.
COMPITO DI COPIA ALLO SPECCHIO
Il soggetto, guardandosi la mano destra riflessa da uno specchio, deve tracciare con una
matita un disegno senza uscire dai margini delle due stelle concentriche. Ogni volta che si
esce dai margini si computa un errore.
La prestazione di H:M: migliora con il tempo e la curva ha lo stesso decorso temporale di
quella ottenuta da un individuo normale.
↓
Memoria procedurale intatta
SISTEMA LIMBICO
Il lobo limbico include la corteccia della parte mediale degli emisferi cerebrali che forma
una sorta di bordo “limbo”, attorno al corpo calloso e al diencefalo e comprende il giro
cingolato e il giro paraippocampale.
IPPOCAMPO
L’ippocampo è una struttura accolta nella compagine della circonvoluzione interna di ogni
lobo temporale. Svolge il ruolo centrale in quella che è la formazione di nuovi ricordi.
Lesioni all'ippocampo causano amnesia anterograda.
Afferenze dell’ippocampo:
· corteccia entorinale
· amigdala
· ippocampo contro laterale
· locus coerulus
Efferenze dell’ippocampo:
Le efferenze si portano soprattutto alle aree associative dei lobi frontale, parietale,
occipitale e temporale. Attraverso il fornice le informazioni provenienti dall’ippocampo
raggiungono i corpi mammillari, il nucleo anteriore e mediodorsale del talamo.
L'ippocampo è solo una parte del circuito della memoria.
Tutta la corteccia è implicata nella memoria
LA PLASTICITÀ NEURONALE
La plasticità neuronale fa sì che “eventi” cerebrali, in risposta a stimoli esterni, possano
determinare modificazioni importanti e persistenti dei circuiti cerebrali. La plasticità
neuronale è alla base dello sviluppo funzionale del SNC, della memoria,
dell’apprendimento.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
esistono due tipi di plasticità sinaptica: presinaptica / postsinaptica
presinaptica: breve termine // postsinaptica: lungo termine
La plasticità sinaptica può essere declinata attraverso il “Principio di Hebb”: se l’assone di
un neurone A è abbastanza vicino per eccitare un altro neurone B in modo ripetuto e
consistente, in uno o in entrambi i neuroni si producono cambiamenti metabolici e un
processo di crescita per cui l’efficienza dei neuroni risulta potenziata.
↳più di neuroni interagiscono tra loro,maggiore sarà la forza della loro interazione e del loro
legame; meno due neuroni interagiscono tra loro, minore sarà la forza della loro interazione
e del loro legame.
>la frequenza di scarica di un neurone, trasmette una parte molto importante di
informazione che riguarda l’aumento di qualcosa.
↪la frequenza di scarica del neurone si la rappresentazione di quella che è l’importanza di
un messaggio neuronale, che altrimenti non potrebbe essere capito in altro modo.
frequenza) è il modo che ha il nostro SNC per dare valore di importanza ad un messaggio
neuronale. Quanto è più alta la frequenza di scarica di un neurone tanto più quel
messaggio ha un qualcosa di importante da dire.
trasmissione del messaggio) arriva u PA nel bottone sinaptico → si aprono i canali del
sodio e poi quelli del potassio → a livello del bottone sinaptico = canali voltaggio
dipendenti si aprono gli altri canali del calcio
→ arriva il PA si aprono i canali del Ca → fenomeno: favorisce la fusione di una vescicola
neurotrasmettitore con la memb. plasmatica del bott. sinaptico
→ neurotrasmettitore liberato → si legherà ad un suo recettore → verrà trasmesso il mess.
PLASTICITÀ A BREVE TERMINE
le vescicole di neurotrasmettitore si accumulano nella parte terminale dell'assone. Il
neurone spara potenziali d’azione, l’impulso elettrico arriva alla terminazione assonale e la
depolarizzazione della membrana provoca l’apertura del canale del calcio che entra
all’interno del bottone assonale.
L’ingresso del calco consente alle vescicole che contengono i neurotrasmettitore di essere
rilasciate nell’area sinaptica. I neurotrasmettitori rilasciati vengono recepiti dai recettori
presenti alla base della cellula postsinaptica.
PLASTICITÀ A LUNGO TERMINE
I recettori del glutammato mediano un fenomeno di plasticità neuronale definito Long Term
Potentiation (LTP), che consiste nel potenziamento, notevole e duraturo, della trasmissione
sinaptica in seguito ad una prima stimolazione “condizionante”, di breve durata ed alta
intensità.
Limite → la p. a lungo termine riguarda solo ed esclusivamente le sinapsi
glutammatergiche.
glutammato: è il principale neurotrasmettitore eccitatorio del SNC, è un costituente delle
proteine. Dividiamo i recettori in due famiglie:
-recettori metabotropici
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
-recettori ionotropici: AMPA / NMDA
Gli astrociti a livello della sinapsi glutammatergica svolgono un ruolo importante. Intorno ai
recettori AMPA/NMDA = gli astrociti è come se sigillassero questo tipo di sinapsi, in
maniera di isolarla dall’esterno.
LA SINAPSI GLUTAMMATERGICA
Recettore AMPA: è un canale ligando dipendente che a seguito del suo legame dal
glutammato fa passare il sodio, è una corrente intensa ma breve. Quando si apre a seguito
del legame con il glutammato permette al sodio di muoversi tra l’esterno verso l'interno.
· Canale per il Na+
· Media affinità per il GLU
Recettore NMDA:
· Permeabile al Ca2+
· Alta affinità per il GLU
· Normalmente bloccato dal Mg2+
· La depolarizzazione della membrana sblocca il canale
L'induzione dell’ LTP richiede l'attività concomitante nel neurone sia presinaptico sia
postsinaptico, secondo il modello proposto da Hebb → Rivelatore di coincidenza
LTP
1. il GLU si lega ai recettori AMPA e NMDA;
2. l’NMDA a riposo è bloccato dal Mg 2+. La membrana
postsinaptica si depolarizza grazie ai flussi di Na+
attraverso gli AMPA;
3. ripetute depolarizzazioni della membrana
postsinaptica rimuovono il blocco del Mg2+. Il Ca2+
entra nel bottone sinaptico via NMDA;
4. il Ca2+ attiva delle proteine chinasi che inducono
modificazioni locali dei bottoni e producono un
messaggero retrogrado (NO?) che, a livello
presinaptico, causa un ulteriore aumento della
liberazione di glutammato;
5. effetto potenziante a feedback positivo che induce
plasticità sinaptica (alterazione funzionale pre- e
post-sinaptica delle sinapsi);
RUOLO DELLA ESPRESSIONE GENICA E DELLA SINTESI PROTEICA NEL
MANTENIMENTO DELL’LTP
La fase finale dell’LTP è dovuta alla PKA che attiva il regolatore trascrizionale CREB che
induce l’espressione di un gruppo di geni che a loro volta producono cambiamenti di lunga
durata nell’attività della PKA e della struttura della sinapsi.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
LONG TERM DEPRESSION (LTD)
1. Concentrazioni di Ca intracellulare più basse e costanti
2. Attivazione di proteinfosfatasi
3. Internalizzazione dei recettori AMPA
4. Diminuzione nella forza di una sinapsi
LONG TERM P/D
Se le sinapsi continuassero a potenziarsi semplicemente in virtù dell’LTP, alla fine
arriverebbero a un punto di massima efficacia e quindi di incapacità di poter codificare
nuove informazioni. Ne deriva che affinché il rafforzamento delle sinapsi possa essere un
processo veramente utile, altri processi come è appunto l’LTD debbano indebolire
selettivamente specifiche sinapsi.
LA PRATICA ALLARGA LA RAPPRESENTAZIONE CORTICALE PREESISTENTE DELLE
AFFERENZE DELLE DITA
Scimmia è addestrata per 1 ora al giorno a eseguire un compito che richiedeva l'uso
ripetuto delle punte del secondo e terzo dito e, occasionalmente anche del quarto. Dopo
l'addestramento, la parte dell'area 3b della corteccia somatosensitiva che rappresenta le
punte delle dita stimolate (colore scuro) è sostanzialmente maggiore del normale (misurata
tre mesi prima dell'addestramento).
Scansione della corteccia motoria, eseguita in fRMI, mostra che dopo tre settimane di
addestramento la regione attivata ne soggetti addestrati (arancione) è maggiore di quella
attivata nei soggetti di controllo che compivano movimenti liberi con la stessa mano. La
variazione nella rappresentazione corticale persisteva, nei soggetti addestrati, per diversi
mesi.
La dimensione della rappresentazione corticale del quinto dito della mano sinistra è
maggiore nei violinisti che in coloro che non suonano uno strumento. Il grafico rappresenta
l'intensità dell'attività neurale. L’aumento è più pronunciato nei musicisti che avevano
iniziato l'addestramento musicale prima di 13 anni. (v.slide 47)
Sinestesia: fenomeno in cui l’esperienza di un senso, ad esempio la vista, ne stimola un
altro, il gusto per esempio.
GLI ERRORI DELLA MEMORIA
· Distrazione: mancanza di attenzione verso le cose che si stanno facendo. Basi neurali
poco conosciute.
· Blocco: incapacità temporanea di ricordare un’informazione conservata in memoria.
Intensa attività della corteccia cingolata anteriore e corteccia prefrontale. Queste attività
sembrano riflettere i tentativi del soggetto di distinguere informazioni sbagliate da quelle
giuste e risolvere il blocco.
· Errata attribuzione: associazione in una memoria con una persona, un luogo e un tempo
errati (falso riconoscimento). Attività dell’ippocampo simili nel corso di riconoscimenti veri
o falsi.
· Distorsione: alterazioni e inserimenti inconsci nella memoria;il passato viene alterato per
renderlo conforme alle esperienze attuali). Basi neurali sconosciute.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
· Suggestionabilità: tendenza a incorporare informazioni estranee nelle nostre memorie
(false memorie possono essere trapiantate in individui particolarmente sensibili attraverso
la ripetizione di episodi che non sono mai avvenuti). Basi neurali sconosciute.
· Persistenza: ricordi ossessivi, costante ricordo di informazioni o eventi che
preferiremmo dimenticare. Può assumere un forte valore adattativo. Forte attivazione
dell’amigdala.
AMNESIA
Condizione patologica caratterizzata dalla perdita di informazioni precedentemente
acquisite e/o dall’incapacità di immagazzinare nuove informazioni:
· anterograda: deficit nell’apprendere nuove informazioni
· retrograda: deficit in richiamare informazioni precedentemente apprese
SISTEMA MOTORIO
I sistemi sensoriali forniscono gli input ai processi cognitivi e i sistemi motori trasmettono
l’output fisico comportamentale che esprime gli scopi cognitivi.
tutti i movimenti corporei sono generati dalla stimolazione delle fibre muscolari scheletriche
da parte dei motoneuroni inferiori, i cui corpi cellulari sono localizzati nel midollo spinale e
cefalico
la corteccia motoria si trova all’interno del lobo frontale ed è costituita da tre aree:
corteccia motoria primaria/ corteccia premotoria/ Area motoria supplementare
controllo motorio
I centri nervosi responsabili del movimento operano su tre livelli:
midollo spinale/sistemi discendenti del tronco-encefalico/ corteccia cerebrale
midollo spinale) distinguiamo due parti: s. bianca - s.grigia
↪nel corno anteriore ci sono i motoneuroni
> nella parte mediale ci sono quelli che innervano i muscoli prossimali, i muscoli distali
↳Nella parte intermedia ci sono gli Inter-neuroni che proiettano a vari livelli dai la stessa
metà del corpo e alla parte controlaterale in modo da regolare la postura
L’attività dei motoneuroni inferiori è coordinata dai interneuroni di circuiti locali; e si sono
regolati da proiezioni discendenti dei motoneuroni superiori della corteccia cerebrale
encefalico.
le aree motorie superiori sono monitorate allora volta da altri due sistemi fondamentali:
→cervelletto: corregge gli errori del movimento in atto e permette di imparare nuove abilità
motorie
→gangli della base: regolano i comandi motori e facilitano forme semplici di apprendimento
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
riflesso= È la risposta involontaria a stimoli di varia natura provenienti dall’ambiente
esterno.
riflesso miotatico> il più semplice di riflessi spinali
-vengono attivati dei recettori = fusi neuromuscolari che trasmettono l’impulso al midollo
spinale, attivando i motoneuroni del quadricipite a contrarsi e quindi il muscolo antagonista
a rilasciarsi.
⤷ I fusi neuromuscolari vengono attivati per via di una percussione avvenuta tramite un
martelletto di gomma sul tendine del quadricipite.
⤷ gamma-motoneuroni modo e livello di attivazione di fusi neuromuscolari contribuendo a
migliorare il controllo sulla lunghezza muscolare.
> movimenti volontari = A differenza dei riflessi i movimenti volontari non sono semplici
risposte a stimoli ambientali ma possono essere generati da programmi motori interni.
vi sono due suddivisioni principali del controllo motorio superiore:
-sistema piramidale: tre fasci cortico-spinali di cui maggiore è il crociato che a livello del
tronco si incrocia e va dall’altra parte del corpo
-sistema extrapiramidale: consiste in diversi componenti separati. Una mia rappresentata
dalla corteccia sensomotoria, sostanza reticolare.
L’altra dalla corteccia motoria, nucleo rosso,. Altre vie mettono in collegamento la corteccia
con i nuclei della base, sostanza reticolare e nucleo rosso.
VIE CORTICOSPINALI E CORTICOBULBARI
i neuroni nella corteccia motoria danno origine ad assoni che viaggiano attraverso la
capsula interna e si fondono sulla superficie ventrale del mesencefalo, all’interno del
peduncolo cerebrale. Questi assoni continuano attraverso i pon e arrivano sulla superficie
ventrale del midollo, dando origine alle piramidi.
↳la maggior parte di queste fibre piramidali si incrociano nella parte caudale del midollo per
formare il tratto corticospinale laterale nella colonna vertebrale.
CORTECCIA MOTORIA PRIMARIA E AREA PREMOTORIA
la corteccia motoria primaria è localizzata nella circonvoluzione precentrale; l’area
premotoria è più rostrale
inizio XX sec:
-Charles Sherrington: pubblica mappe motorie sulle scimmie antropomorfe usando la
stimolazione elettrica focale.
-Wilder Penfield: dimostra la mappa muscolotopica stimolando la corteccia motoria umana.
CORTECCIA MOTORIA PRIMARIA
possiede una apa chiamata mappa somatotopica; cioè ogni pezzo della corteccia motoria
primaria rappresenta una parte del corpo.
↓
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
area motrice primaria: contiene una mappa motoria del corpo homunculus. L‘attivazione
(area 4 di Brodmann) è collegata principalmente a movimenti semplici di singole
articolazioni
Precisi schemi di movimento vengono comandati da popolazioni di motoneuroni corticali
sintonizzati tra loro.
DIVISIONE DELLA C.MOTORIA NEL CERVELLO DI UN MACACO
come negli esseri umani la corteccia motoria primaria si trova nel bordo anteriore del solo
centrale. Ventralmente a questa regione, un complesso di aree premotorie si estende
dell'opercolo frontale della superficie posteriore del lobo frontale alla circonvoluzione
cingolgolata sulla superficie dell'emisfero mediale.
Le aree premotorie laterali e le aree motorie supplementari sono dedicate alla selezione e
all’organizzazione di movimenti preciso del viso e delle mani
⤷i campi oculari frontali organizzano il movimento volontario dello sguardo; le aree motorie
del cingolo sono coinvolte nell’espressione del comportamento emozionale somatico.
CORTECCIA PREMOTORIA
I neuroni della corteccia premotoria influiscono sul comportamento motorio:
-indirettamente → estese connessioni reciproche con M1
-direttamente → attraverso fibre che scendono nel fascio corticospinale
l’esistenza di connessioni dirette mette in dubbio l’ipotesi che la corteccia premotoria sia
gerarchicamente superiore alla corteccia motoria primaria.
>utilizza le informazioni provenienti da altre aree corticali per selezionare elementi adatti al
contesto dell’azione —> scelta dell’azione
-area p. Laterale: scelta sulla base degli elementi esterni
-area p. Mediale: scelta sulla base degli elementi interni
>codifica l’intenzione dell’azione
>codificano per movimenti osservati piuttosto che eseguiti = neuronispecchio
AREE PREMOTORIE MEDIALI (supplementare e pre-supplementare)
Implicate nella definizione dei programmi motori e nel loro apprendimento.
Quando appreso il controllo può passare dalla supplementare alla motrice primaria
Esecuzione semplice: si attivano area motoria e somatosensitiva
Esecuzione complessa: si attivano area motoria, somatosensitiva e supplementare
AREE PREMOTORIE LATERALI
Sono implicate nell’associazione di eventi sensoriali con particolari movimenti e controllano
i movimenti guidati da informazioni visive. Durante i movimenti complessi i neuroni dell’area
parietale 5 potrebbero ricevere informazioni dei movimenti nel corso del movimento per
operare un controllo dei movimenti da parte delle aree premotorie.
NEURONI SPECCHIO
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Si attivano sia quando si sta eseguendo un determinato movimento, ma si attivano anche
quando qualcun altro sta eseguendo quello stesso movimento.
⤷potrebbero andare a comprendere le azioni altrui
GANGLI DELLA BASE
Sono: => sono circuiti diretti e indiretti e partono dalla
-striato (nucleo caudato + putamen) Corteccia per poi arrivare a diverse aree
-globus pallidus Corticali
-nucleo subtalamico
-sostanza nera
Via diretta: attiva il movimento
Via indiretta:inibisce il movimento
-i gangli della base forniscono un ponte tra corteccia cerebrale e talamo con ritorno alla
corteccia. Tutto questo sistema è deputato alla regolazione del movimento. I gangli della
base non generano il movimento, essi si attivano solo a movimento iniziato per controllarlo
e regolarlo; identificano l’obiettivo del movimento nello spazio.
>il talamo manda connessioni eccitatorie alla corteccia per far avvenire le azioni. Il nucleo
GPi di base inibisce sempre il talamo; lo striato riceve connessioni eccitatorie dalla
corteccia e a sua volta va ad inibire lo GPi.
↳se l GPi è inibito non esiste più l’inibizione sul talamo.
>il GPe inibisce il GPi, che a sua volta è inibito dallo striato. L’azione inibitoria del GPe sul
GPi non esiste più.
↳il nucleo subtalamico è eccitato dalla corteccia ed eccita il GPi. Di base il GP inibisce il
nucleo subtalamico.
La sostanza nera è importantissima; facilita la via diretta e inibisce la via indiretta.
Quando i suoi neuroni degenerano/si danneggiano, le sue azioni non sono più facilitate e
quindi vi è un deficit di movimento.
MOTRICITÀ CORTICALE
La corteccia motrice controlla i motoneuroni spinali direttamente attraverso il fascio
cortico-spinale laterale (via piramidale) e indirettamente attraverso vie che originano dai
centri motori del tronco encefalico (via non piramidali).
Area motoria secondaria:
-stimolazione: inibizione dei movimenti in corso o comparsa del desiderio di compiere un
movimento.
Area motoria supplementare:
-rappresenta un’estensione dell’area 6 verso la faccia interna, inter-emisferica, della
corteccia cerebrale.
-stimolazione: movimenti complessi o componente toracica.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
RUOLO DELL’AREA 6
É costituita dalla corteccia supplementare motoria (mediale) e dalla corteccia premotoria
(laterale). L’area ha una posizione gerarchicamente superiore all’area 4, sarebbe cioè
analoga ad un’area associativa motoria.
RUOLO FUNZIONALE DELLA VIA PIRAMIDALE
Alterazione della motricità fine, soprattutto a carico della mano. I movimenti sono rallentati,
l'afferramento degli oggetti è possibile solo con la chiusura simultanea di tutte le dita.
La motricità fine di precisione sia dovuta alle connessioni monosinaptiche fra neurone
corticale e motoneurone spinale. Il fascio piramidale raggiunge la sua massima
espressione evolutiva nei primati e soprattutto nell’uomo dove influenza soprattutto i
muscoli distali, ciò sottolinea l’enorme importanza di questo sistema per le abilità manuali
dell’uomo.
SISTEMA EXTRAPIRAMIDALE
È costituito da numerose strutture anatomiche e vie motorie che hanno un ruolo
fondamentale: nell’iiniziativa motoria/ nell’esercitazione di gesti ritmici/ mantenimento della
postura
FORMAZIONE RETICOLARE
La posizione della formazione reticolare In relazione ad altri principali punti di riferimento a
diversi livelli del tronco encefalico i neuroni Nella formazione reticolare sono sparsi tra i
fasci di assoni che passano attraverso la parte mediale del pon, il mesencefalo, il midollo.
Document shared on https://www.docsity.com/it/appunti-di-neurofisiologia-9/8617984/
Downloaded by: srd30.01 (draimond.sara@gmail.com)
Potrebbero piacerti anche
- Tecniche Di Massaggio SportivoDocumento49 pagineTecniche Di Massaggio Sportivogigi_sausage100% (5)
- Sistema NervosoDocumento26 pagineSistema NervosoTessy PsibarNessuna valutazione finora
- Alias Supplemento Del Manifesto 02/07/2011Documento24 pagineAlias Supplemento Del Manifesto 02/07/2011harck181Nessuna valutazione finora
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- Il Sistema NervosoDocumento6 pagineIl Sistema NervosoMartina MusmeciNessuna valutazione finora
- Conduzione Impulso NervosoDocumento32 pagineConduzione Impulso NervosoMarcoNessuna valutazione finora
- L'impulso NervosoDocumento7 pagineL'impulso NervosoGiancarlo AttivissimoNessuna valutazione finora
- Dispensa Fisiologia UnitoDocumento497 pagineDispensa Fisiologia Unitodorian88Nessuna valutazione finora
- Il Sistema NervosoDocumento8 pagineIl Sistema NervosoMartina RossiNessuna valutazione finora
- Sinapsi e NeurotrasmettitoriDocumento45 pagineSinapsi e NeurotrasmettitoriNicola Laurora0% (1)
- FISIOLOGIAsbobineDocumento164 pagineFISIOLOGIAsbobinemarco.ricco.mr00Nessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- Appunti NeurofisiologiaDocumento39 pagineAppunti NeurofisiologiaGianluca LibianiNessuna valutazione finora
- Conduzione NervosaDocumento10 pagineConduzione NervosaMiriam LongobardiNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- CITOLOGIA E ISTOLOGIA - Lezione 7Documento2 pagineCITOLOGIA E ISTOLOGIA - Lezione 7Federico TosiniNessuna valutazione finora
- Fisiologia SinapsiDocumento1 paginaFisiologia SinapsiFedericaRomanoNessuna valutazione finora
- NeurofisiologiaDocumento69 pagineNeurofisiologiafabiana900% (1)
- (Medicina-Ita) Neurologia - EEGDocumento44 pagine(Medicina-Ita) Neurologia - EEGedosuper1Nessuna valutazione finora
- FisiologiaDocumento42 pagineFisiologiaLaura TreggiariNessuna valutazione finora
- Fisiologia ValenteDocumento4 pagineFisiologia ValenteFrancesco FalconieriNessuna valutazione finora
- Scienze - Il Sistema NervosoDocumento27 pagineScienze - Il Sistema NervosoRiccardo PengoNessuna valutazione finora
- Tessuto NervosoDocumento7 pagineTessuto NervosoeristelaNessuna valutazione finora
- Sistema NervosoDocumento60 pagineSistema NervosoAnnapaola PoliNessuna valutazione finora
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- BiologiaDocumento6 pagineBiologiaracheleNessuna valutazione finora
- NEUROBIOLOGIA La SapienzaDocumento30 pagineNEUROBIOLOGIA La Sapienzachiara cimeiNessuna valutazione finora
- Anche Ad Altre ComponentiDocumento4 pagineAnche Ad Altre Componentifrancesco biancoNessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Sistema NervosoDocumento15 pagineSistema NervosoBruce WiiNessuna valutazione finora
- Fondamenti Anatomo Fisiologici Dellattivita PsichicaDocumento29 pagineFondamenti Anatomo Fisiologici Dellattivita PsichicagigioNessuna valutazione finora
- Docsity Domande Triennale Fisiologia UmanaDocumento33 pagineDocsity Domande Triennale Fisiologia Umanadestinyema78Nessuna valutazione finora
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- Fisiologia Cellulare Potenziale AzioneDocumento31 pagineFisiologia Cellulare Potenziale Azionesimo_pNessuna valutazione finora
- Documento Senza TitoloDocumento15 pagineDocumento Senza TitoloCarlotta MazzoniNessuna valutazione finora
- Marieb Sintesi 62091 c14Documento3 pagineMarieb Sintesi 62091 c14Caterina NuzzoNessuna valutazione finora
- Biologia 2Documento75 pagineBiologia 2Sara DedolaNessuna valutazione finora
- FISIOLOGÍADocumento26 pagineFISIOLOGÍAAndrea De La FuenteNessuna valutazione finora
- NeurologiaDocumento50 pagineNeurologiaSalvatore FerraroNessuna valutazione finora
- Conduzione Nervosa (Riassunto)Documento10 pagineConduzione Nervosa (Riassunto)Aaron RomanoNessuna valutazione finora
- Il Sistema NervosoDocumento14 pagineIl Sistema NervosoMartina BoriaNessuna valutazione finora
- Fisiologia Lezione 4Documento6 pagineFisiologia Lezione 4The last chimeraNessuna valutazione finora
- 1 Potenziale Di Equilibrio Di NernstDocumento5 pagine1 Potenziale Di Equilibrio Di NernstClaudia OrlannoNessuna valutazione finora
- Trasmissione Sinaptica e Integrazione NeuronaleDocumento11 pagineTrasmissione Sinaptica e Integrazione NeuronaleAnnamaria MaderaNessuna valutazione finora
- Sistema Nervoso - 24 Maggio 2023Documento34 pagineSistema Nervoso - 24 Maggio 2023Gianluca TortoraNessuna valutazione finora
- 10 - Potenziali Graduati e Potenziale D-AzioneDocumento48 pagine10 - Potenziali Graduati e Potenziale D-AzioneMarcoNessuna valutazione finora
- 0011 - Appunti Di FisiologiaDocumento0 pagine0011 - Appunti Di FisiologiaRoberta ConennaNessuna valutazione finora
- Potenziali d'AzioneDocumento5 paginePotenziali d'AzioneHghh NgthhNessuna valutazione finora
- Appunti Fisiologia-2Documento39 pagineAppunti Fisiologia-2Serena D'AmicoNessuna valutazione finora
- Sistema NervosoDocumento6 pagineSistema NervosoGraziella RosanoNessuna valutazione finora
- Il Sistema NervosoDocumento4 pagineIl Sistema NervosoGiulia ConiglioNessuna valutazione finora
- Anatomia Del Sistema NervosoDocumento16 pagineAnatomia Del Sistema NervosoAlysiaNessuna valutazione finora
- 0011 - Appunti Di FisiologiaDocumento152 pagine0011 - Appunti Di Fisiologiasilasjuba100% (1)
- Domande e Risposte Sistema NervosoDocumento4 pagineDomande e Risposte Sistema Nervosoedup220% (1)
- 4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azioneDocumento43 pagine4) Il Sistema Nervoso. Le Cellule Del Sistema Nervoso. Il Potenziale D'azionedavide di lorenzoNessuna valutazione finora
- Tessuto NervosoDocumento6 pagineTessuto NervosoDinjo79Nessuna valutazione finora
- Sbobine Fisiologia Umana I 2015Documento443 pagineSbobine Fisiologia Umana I 2015Leonardo MarchettiNessuna valutazione finora
- Cristal 6Documento32 pagineCristal 6Vale46 RossiNessuna valutazione finora
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- Le Catene Lineari del Corpo davanti alla MetapsicologiaDa EverandLe Catene Lineari del Corpo davanti alla MetapsicologiaNessuna valutazione finora
- FisiologiaDocumento429 pagineFisiologiaAndrea Di GiorgioNessuna valutazione finora
- Spezifikation SonoScape S40Documento18 pagineSpezifikation SonoScape S40Gomez, Francisco (AJR)Nessuna valutazione finora
- Ix 2013 X - Xi - Xii PDFDocumento43 pagineIx 2013 X - Xi - Xii PDFGiuseppe FoggiaNessuna valutazione finora
- Ursula K. Le Guin - Città Delle IllusioniDocumento154 pagineUrsula K. Le Guin - Città Delle Illusioninikle3Nessuna valutazione finora
- The SauraDocumento28 pagineThe SauraioginevraNessuna valutazione finora