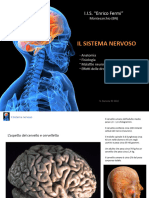
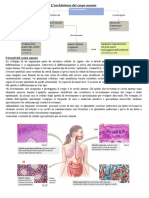
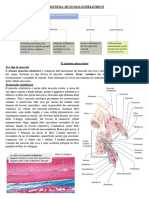
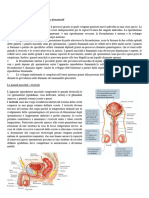
Potrebbero piacerti anche
- Riassunto Bear Neuroscienze Esplorando Il CervelloDocumento9 pagineRiassunto Bear Neuroscienze Esplorando Il CervellocdeipoliNessuna valutazione finora
- Manuale Empire EarthDocumento238 pagineManuale Empire Earthettore100% (1)
- Il Sistema NervosoDocumento14 pagineIl Sistema NervosoMartina BoriaNessuna valutazione finora
- Il Sistema NervosoDocumento4 pagineIl Sistema NervosoGiulia ConiglioNessuna valutazione finora
- Sistema NervosoDocumento60 pagineSistema NervosoAnnapaola PoliNessuna valutazione finora
- NeurologiaDocumento50 pagineNeurologiaSalvatore FerraroNessuna valutazione finora
- Fisiologia Del ComportamentoDocumento20 pagineFisiologia Del ComportamentoracheleNessuna valutazione finora
- Docsity Riassunto Neuropsicologia Universita MercatorumDocumento81 pagineDocsity Riassunto Neuropsicologia Universita Mercatorumpugliesegiuseppe229Nessuna valutazione finora
- Riassunto Neuropsicologia 01 11 2023Documento158 pagineRiassunto Neuropsicologia 01 11 2023pugliesegiuseppe229Nessuna valutazione finora
- LE BASI PSICOFISIOLOGICHE DEL COMPORTAMENTO DocDocumento14 pagineLE BASI PSICOFISIOLOGICHE DEL COMPORTAMENTO DocElena CampaniNessuna valutazione finora
- Tessuto NervosoDocumento14 pagineTessuto NervosocNessuna valutazione finora
- Appunti Fisiologia-2Documento39 pagineAppunti Fisiologia-2Serena D'AmicoNessuna valutazione finora
- Sistema Nervoso, Il Tessuto NervosoDocumento3 pagineSistema Nervoso, Il Tessuto NervosoNahomie DonfackNessuna valutazione finora
- Domande e Risposte Sistema NervosoDocumento4 pagineDomande e Risposte Sistema Nervosoedup220% (1)
- Scienze BiomedicheDocumento21 pagineScienze BiomedicheRenato RossiNessuna valutazione finora
- Sistema NervosoDocumento34 pagineSistema Nervosomarkcost100% (2)
- Tessuto NervosoDocumento7 pagineTessuto NervosoeristelaNessuna valutazione finora
- 05 Sistema NervosoDocumento12 pagine05 Sistema Nervososono bruttoNessuna valutazione finora
- PSICOLOGIA: La Conduzione NeuronaleDocumento1 paginaPSICOLOGIA: La Conduzione NeuronaleElena della VellaNessuna valutazione finora
- Anatomia Del Sistema NervosoDocumento57 pagineAnatomia Del Sistema NervosoGiorgio MarrasNessuna valutazione finora
- Appunti NeurofisiologiaDocumento39 pagineAppunti NeurofisiologiaGianluca LibianiNessuna valutazione finora
- Dispensa Acustica CervelloDocumento20 pagineDispensa Acustica CervelloAlfonso BelfioreNessuna valutazione finora
- Il Sistema Nervoso SemplificatoDocumento5 pagineIl Sistema Nervoso SemplificatoFrancescaNessuna valutazione finora
- Fondamenti Anatomo Fisiologici Dellattivita PsichicaDocumento29 pagineFondamenti Anatomo Fisiologici Dellattivita PsichicagigioNessuna valutazione finora
- Scienze - Il Sistema NervosoDocumento27 pagineScienze - Il Sistema NervosoRiccardo PengoNessuna valutazione finora
- Il Sistema NervosoDocumento5 pagineIl Sistema NervosoGianluca LammendolaNessuna valutazione finora
- Sistema NervosoDocumento31 pagineSistema NervosoNicola RainoneNessuna valutazione finora
- Biologia 2Documento75 pagineBiologia 2Sara DedolaNessuna valutazione finora
- Fisiologia 4 - La Comunicazione NervosaDocumento25 pagineFisiologia 4 - La Comunicazione NervosaAnna De Simone100% (1)
- Anatomia Sistema Nervoso PDFDocumento19 pagineAnatomia Sistema Nervoso PDFSimone EspositoNessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- Anatomia e Fisopatologia 2021Documento137 pagineAnatomia e Fisopatologia 2021Ejona SabijaNessuna valutazione finora
- Aula Sistema NervosoDocumento34 pagineAula Sistema NervosoluizvitalcNessuna valutazione finora
- Sistema Nervoso Centrale 1Documento14 pagineSistema Nervoso Centrale 1Miriam LongobardiNessuna valutazione finora
- Il Sistema NervosoDocumento7 pagineIl Sistema NervosoEmail SitiNessuna valutazione finora
- FISIOLOGIADocumento6 pagineFISIOLOGIAMariateresa SantorsolaNessuna valutazione finora
- Il Sistema NervosoDocumento3 pagineIl Sistema NervosoMatteo QuintilianiNessuna valutazione finora
- Anatomia Del Sistema NervosoDocumento16 pagineAnatomia Del Sistema NervosoAlysiaNessuna valutazione finora
- Dispensa NeurologiaDocumento14 pagineDispensa NeurologiaDaniele SicilianoNessuna valutazione finora
- Sistema NervosoDocumento23 pagineSistema NervosoDiana ChiccaNessuna valutazione finora
- Principi NeuroscienzeDocumento12 paginePrincipi NeuroscienzegwalidorNessuna valutazione finora
- Tessuto Nervoso: Suddivisione Anatomica Generale Prevede UnDocumento10 pagineTessuto Nervoso: Suddivisione Anatomica Generale Prevede UnLuana LeonardiNessuna valutazione finora
- BiologiaDocumento6 pagineBiologiaracheleNessuna valutazione finora
- PW 2Documento4 paginePW 2GiorgioNessuna valutazione finora
- Appunti NeuroscienzeDocumento131 pagineAppunti NeuroscienzeDavide ValentiNessuna valutazione finora
- Presentazione Sistema NervosoDocumento60 paginePresentazione Sistema NervosoMattNessuna valutazione finora
- Sistema NervosoDocumento6 pagineSistema NervosoGraziella RosanoNessuna valutazione finora
- 1 NeuroniDocumento10 pagine1 NeuronifNessuna valutazione finora
- Marieb Sintesi 62091 c14Documento3 pagineMarieb Sintesi 62091 c14Caterina NuzzoNessuna valutazione finora
- Sistema Nervoso RiassuntoDocumento64 pagineSistema Nervoso Riassuntolucchini.paolo3002Nessuna valutazione finora
- Sistema Nervoso - 24 Maggio 2023Documento34 pagineSistema Nervoso - 24 Maggio 2023Gianluca TortoraNessuna valutazione finora
- Conduzione Impulso NervosoDocumento32 pagineConduzione Impulso NervosoMarcoNessuna valutazione finora
- 11-Sistema Nervoso 2017Documento91 pagine11-Sistema Nervoso 2017Murilo AlbuquerqueNessuna valutazione finora
- Copia Di Copia Di Neuropedagogia Infantile Completa 2Documento106 pagineCopia Di Copia Di Neuropedagogia Infantile Completa 2Dafne PanebiancoNessuna valutazione finora
- Tecniche della riabilitazione graficaDocumento27 pagineTecniche della riabilitazione graficagiorgia.sacchieroNessuna valutazione finora
- Dispensa NeuroanatomiaDocumento142 pagineDispensa NeuroanatomiaPierpaolo LuzioNessuna valutazione finora
- Sistema NervosoDocumento5 pagineSistema NervosoGiuliaNessuna valutazione finora
- Riassunto Neuro ScienzeDocumento25 pagineRiassunto Neuro Scienzeflavia01cubisinoNessuna valutazione finora
- Sistema NervosoDocumento3 pagineSistema NervosoEleonora RivaNessuna valutazione finora
- Nervo Vago: La Guida Completa per Scoprire come Attivare i Benefici Curativi del tuo Nervo Vago – Riduci con Esercizi di Self-Help l’Ansia, Depressione, Infiammazioni, Rabbia e TraumiDa EverandNervo Vago: La Guida Completa per Scoprire come Attivare i Benefici Curativi del tuo Nervo Vago – Riduci con Esercizi di Self-Help l’Ansia, Depressione, Infiammazioni, Rabbia e TraumiNessuna valutazione finora
- L'OMEOSTASIDocumento4 pagineL'OMEOSTASICarlotta MazzoniNessuna valutazione finora
- BIOLOGIADocumento6 pagineBIOLOGIACarlotta MazzoniNessuna valutazione finora
- Il Sistema Muscolo-ScheletricoDocumento4 pagineIl Sistema Muscolo-ScheletricoCarlotta MazzoniNessuna valutazione finora
- L'Apparato RiproduttoreDocumento14 pagineL'Apparato RiproduttoreCarlotta MazzoniNessuna valutazione finora
- BIOLOGIADocumento6 pagineBIOLOGIACarlotta MazzoniNessuna valutazione finora
- Il Sistema Muscolo-ScheletricoDocumento4 pagineIl Sistema Muscolo-ScheletricoCarlotta MazzoniNessuna valutazione finora
- Il Sistema PeriodicoDocumento9 pagineIl Sistema PeriodicoCarlotta MazzoniNessuna valutazione finora
- Inno Del Primo MaggioDocumento1 paginaInno Del Primo MaggioGabriele torconeNessuna valutazione finora
- XYZ Degli OscilloscopiDocumento64 pagineXYZ Degli OscilloscopiPierluigi PollanoNessuna valutazione finora
- Dermal Fillers Di Acido Ialuronico PDFDocumento4 pagineDermal Fillers Di Acido Ialuronico PDFPaoly Palma100% (1)
- Come Si Usa Il TesterDocumento13 pagineCome Si Usa Il TesterJackNessuna valutazione finora
- Imperial Academy of RussiaDocumento4 pagineImperial Academy of RussiaEmanuele NuscaNessuna valutazione finora
- La Ricerca Dell'ImmortalitàDocumento6 pagineLa Ricerca Dell'ImmortalitàclaudioNessuna valutazione finora
- Analisi Critica Delle OrgonitiDocumento19 pagineAnalisi Critica Delle OrgonitiD.Domenico100% (1)