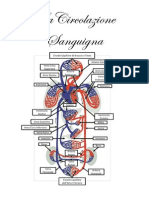
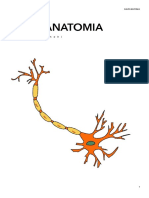
Potrebbero piacerti anche
- Forme degli Animali VERTEBRATI: Caratteri Fondamenta e Forme AdattativeDa EverandForme degli Animali VERTEBRATI: Caratteri Fondamenta e Forme AdattativeNessuna valutazione finora
- Forme degli Animali INVERTEBRATI: Classificazione e Formazione dei Corpi AnimaliDa EverandForme degli Animali INVERTEBRATI: Classificazione e Formazione dei Corpi AnimaliNessuna valutazione finora
- CiclostomiDocumento12 pagineCiclostomiManuela BolognaNessuna valutazione finora
- 2 - Esame Scritto Di Zoologia Dei Vertebrati Pt.1Documento16 pagine2 - Esame Scritto Di Zoologia Dei Vertebrati Pt.1molesdan3540Nessuna valutazione finora
- RettiliDocumento14 pagineRettiliManuela BolognaNessuna valutazione finora
- Liquido Amniotico, Ma in Un Uovo Che Può Essere Deposto Sulla Terra Emersa. I Primi Sono I Rettili CheDocumento10 pagineLiquido Amniotico, Ma in Un Uovo Che Può Essere Deposto Sulla Terra Emersa. I Primi Sono I Rettili CheWendy Marilyn Ysamar AlvaroNessuna valutazione finora
- I RettiliDocumento3 pagineI RettiliEdoardo MigliavaccaNessuna valutazione finora
- Ex Malacostraca (Copepodi, Branchiuri, Cirripedi, Ostracodi)Documento7 pagineEx Malacostraca (Copepodi, Branchiuri, Cirripedi, Ostracodi)Dottor JergalNessuna valutazione finora
- 5 - Classe AnfibiDocumento82 pagine5 - Classe AnfibiEugenio MolestiNessuna valutazione finora
- CephalopodaDocumento4 pagineCephalopodagmc.messanaNessuna valutazione finora
- Anatomia Comparata SbobDocumento52 pagineAnatomia Comparata SbobthemirroNessuna valutazione finora
- Domande Di Filogenesi Animale PDFDocumento6 pagineDomande Di Filogenesi Animale PDFMarina TigrisNessuna valutazione finora
- 3 - Classificazione Dei Vertebrati pt.3 (Riassunti)Documento29 pagine3 - Classificazione Dei Vertebrati pt.3 (Riassunti)molesdan3540Nessuna valutazione finora
- 8 Anatomia Comparata Appunti CompletiDocumento98 pagine8 Anatomia Comparata Appunti CompletiEugenio Molesti100% (1)
- 02coralli Gasteropodi BivalviDocumento7 pagine02coralli Gasteropodi BivalvipplotticiNessuna valutazione finora
- I CheloniDocumento4 pagineI CheloniEdoardo MigliavaccaNessuna valutazione finora
- Appunti Zoologia Dei VertebratiDocumento40 pagineAppunti Zoologia Dei Vertebratidsofia2001Nessuna valutazione finora
- AnfibiDocumento16 pagineAnfibiManuela BolognaNessuna valutazione finora
- Crostacei DecapodiDocumento8 pagineCrostacei DecapodiDottor JergalNessuna valutazione finora
- 1 - Classificazione Dei VertebratiDocumento149 pagine1 - Classificazione Dei VertebratiEugenio MolestiNessuna valutazione finora
- Mollusc ADocumento9 pagineMollusc Agmc.messanaNessuna valutazione finora
- Gli Animali Vertebrati-SDocumento23 pagineGli Animali Vertebrati-Snomoreheroes161Nessuna valutazione finora
- Sistematica Dei Crostacei DecapodiDocumento4 pagineSistematica Dei Crostacei DecapodiElden ring preorderNessuna valutazione finora
- ZoologiaDocumento4 pagineZoologiagiamatteo89Nessuna valutazione finora
- Descrizione Scientifica Degli AnimaliDocumento2 pagineDescrizione Scientifica Degli AnimaliMassimo DeplanoNessuna valutazione finora
- 8 Molluschi 2014Documento65 pagine8 Molluschi 2014Adriana Pellitteri100% (1)
- Lavoro Invertebrati ICDocumento26 pagineLavoro Invertebrati ICnomoreheroes161Nessuna valutazione finora
- AnatomiaComparataLezione3 Dic 2013Documento6 pagineAnatomiaComparataLezione3 Dic 2013Antonio De GaetanoNessuna valutazione finora
- 11 Radiolari HRDocumento6 pagine11 Radiolari HRrazta35Nessuna valutazione finora
- Anatomia Comparata - Storia Della Vita e Degli Animali PDFDocumento149 pagineAnatomia Comparata - Storia Della Vita e Degli Animali PDFNannai02Nessuna valutazione finora
- 10 Anatomia Pt.5Documento81 pagine10 Anatomia Pt.5Eugenio MolestiNessuna valutazione finora
- PhylumDocumento14 paginePhylumWendy Marilyn Ysamar AlvaroNessuna valutazione finora
- Ungulati: Biologia e GestioneDocumento50 pagineUngulati: Biologia e GestionedebugsterNessuna valutazione finora
- Zoo AperteeeeDocumento4 pagineZoo Aperteeeefocaxo5386Nessuna valutazione finora
- Gli AracnidiDocumento21 pagineGli Aracnidimpaola68Nessuna valutazione finora
- PESCA A MOSCA - Entomologia BaseDocumento17 paginePESCA A MOSCA - Entomologia BaseLopez Ivan100% (1)
- Anatomia ComparataDocumento106 pagineAnatomia ComparataAndrea GattoNessuna valutazione finora
- Gli AnimaliDocumento63 pagineGli AnimaliNinna NannaNessuna valutazione finora
- 06 MOLLUSCHI e SIPUNCULIDocumento27 pagine06 MOLLUSCHI e SIPUNCULIEugenio StracuzzaNessuna valutazione finora
- 04 PlatelmintiDocumento17 pagine04 PlatelmintiEugenio StracuzzaNessuna valutazione finora
- Zoologia SistematicaDocumento67 pagineZoologia SistematicaDavide LoiNessuna valutazione finora
- I MammiferiDocumento11 pagineI MammiferiRaffaello PrandoNessuna valutazione finora
- 03 Cnidari e CtenoforiDocumento25 pagine03 Cnidari e CtenoforiEugenio StracuzzaNessuna valutazione finora
- 10 Euartropodi Ii CrostaceiDocumento39 pagine10 Euartropodi Ii CrostaceiEugenio StracuzzaNessuna valutazione finora
- Hordati Parc 2Documento21 pagineHordati Parc 2Lejla MujakićNessuna valutazione finora
- RESPIRATORIODocumento4 pagineRESPIRATORIOMarta OreficiniNessuna valutazione finora
- PR Berlese 1909 MyrientomataDocumento182 paginePR Berlese 1909 MyrientomataCarlo JacominiNessuna valutazione finora
- 02 Poriferi e PlacozoiDocumento5 pagine02 Poriferi e PlacozoiEugenio StracuzzaNessuna valutazione finora
- Appunti Entomologia 1Documento13 pagineAppunti Entomologia 1Fabio LandornoNessuna valutazione finora
- Anatomia ComparataDocumento43 pagineAnatomia ComparataVeronica ViolaNessuna valutazione finora
- Classificazione Organismi MariniDocumento19 pagineClassificazione Organismi MariniterezaNessuna valutazione finora
- Biologia Dei MammiferiDocumento49 pagineBiologia Dei MammiferiLucaCangialosiNessuna valutazione finora
- Appunti Di ZoologiaDocumento123 pagineAppunti Di ZoologiaDario Marchese100% (1)
- RITROVAMENTO DI RESTI SCHELETRICI DI UN DElFINO NEL PliOCENE MEDIO-SUPERIORE DEI DINTORNI DI SETTIME (ASTI, ITALIA NW) .Documento15 pagineRITROVAMENTO DI RESTI SCHELETRICI DI UN DElFINO NEL PliOCENE MEDIO-SUPERIORE DEI DINTORNI DI SETTIME (ASTI, ITALIA NW) .Gianluca BoninsegniNessuna valutazione finora
- Ecologia Marina, Massimo Ponti, Università Di BolognaDocumento33 pagineEcologia Marina, Massimo Ponti, Università Di BolognaSofia PieriNessuna valutazione finora
- Biologia Dello SviluppoDocumento12 pagineBiologia Dello SviluppoManuela BolognaNessuna valutazione finora
- Presentazione 6Documento7 paginePresentazione 6Diego MeatuNessuna valutazione finora
- Contributo alla conoscenza della Malacofauna Marina della Riviera del Conero e dintorniDa EverandContributo alla conoscenza della Malacofauna Marina della Riviera del Conero e dintorniNessuna valutazione finora
- ANATOMIA NoraDocumento4 pagineANATOMIA NoradanixttNessuna valutazione finora
- I VertebratiDocumento3 pagineI VertebratimariaNessuna valutazione finora
- I Primi Passi Verso La Ricchezza Vera 2Documento68 pagineI Primi Passi Verso La Ricchezza Vera 2Manuela BolognaNessuna valutazione finora
- I Primi Passi Verso La Ricchezza Vera 1Documento31 pagineI Primi Passi Verso La Ricchezza Vera 1Manuela BolognaNessuna valutazione finora
- FagaceeDocumento1 paginaFagaceeManuela BolognaNessuna valutazione finora
- Mammiferi Parte 1Documento16 pagineMammiferi Parte 1Manuela BolognaNessuna valutazione finora
- AnfibiDocumento16 pagineAnfibiManuela BolognaNessuna valutazione finora
- Biologia Dello SviluppoDocumento12 pagineBiologia Dello SviluppoManuela BolognaNessuna valutazione finora
- Lezione 03 - Neurologia - 2021-2022 Semeiologia Funzione Motoria Gangli Della Base Coordinazione. Funzione SensitivaDocumento124 pagineLezione 03 - Neurologia - 2021-2022 Semeiologia Funzione Motoria Gangli Della Base Coordinazione. Funzione Sensitivamaria cristina ristagnoNessuna valutazione finora
- FisiologiaDocumento4 pagineFisiologiaHghh NgthhNessuna valutazione finora
- 1° Corso - Osteopatia Craniale Vol 1 - Le Disfunzioni Della PDFDocumento89 pagine1° Corso - Osteopatia Craniale Vol 1 - Le Disfunzioni Della PDFpaolo68audisio100% (3)
- TTD - TMU - Lezioni I - XIIDocumento81 pagineTTD - TMU - Lezioni I - XIIMaegazine EnizageamNessuna valutazione finora
- CircolatorioDocumento41 pagineCircolatorioRiccardo Mason TroiaNessuna valutazione finora
- La Biologia Delle Emozioni 9788896865422 637111Documento225 pagineLa Biologia Delle Emozioni 9788896865422 637111Elisa Osimani100% (4)
- NeuroDocumento9 pagineNeuroMatteo FerrariNessuna valutazione finora
- Anatomia Del Sistema Nervoso Centrale - Schemi VisiviDocumento58 pagineAnatomia Del Sistema Nervoso Centrale - Schemi VisiviClara ScopellitiNessuna valutazione finora
- 29 BisDocumento41 pagine29 Bissupernatura100% (5)
- Anatomia II Medicina PisaDocumento100 pagineAnatomia II Medicina PisanuunumclNessuna valutazione finora
- Appunti NeruopsicologiaDocumento32 pagineAppunti NeruopsicologiaMartina CalvoNessuna valutazione finora
- Le 5 Leggi Di Hamer-Medicina AlternativaDocumento4 pagineLe 5 Leggi Di Hamer-Medicina AlternativaVico Massagista e Quiropraxia - Massagem, Massoterapia e Ventosa Terapia - São José SC.100% (1)
- Sistema Nervoso Encefalo e Nervi Cranici Capitolo 15Documento27 pagineSistema Nervoso Encefalo e Nervi Cranici Capitolo 15enrico73Nessuna valutazione finora
- Tabella Vie SNCDocumento3 pagineTabella Vie SNCLeonardo Pzl100% (1)
- Neoplasie CerebraliDocumento12 pagineNeoplasie CerebraliPatrizia CastellanetaNessuna valutazione finora
- Sistema Nervoso RiassuntoDocumento64 pagineSistema Nervoso Riassuntolucchini.paolo3002Nessuna valutazione finora
- Marchetti M., Pillastrini P. Neurofisiologia Del MovimentoDocumento550 pagineMarchetti M., Pillastrini P. Neurofisiologia Del MovimentoAttilio LiberatoNessuna valutazione finora
- Lezione 1 NeuroradiologiaDocumento13 pagineLezione 1 NeuroradiologiaFraNessuna valutazione finora
- GRAFOLOGIADocumento4 pagineGRAFOLOGIAJoePansaNessuna valutazione finora
- Psicologia MusicaleDocumento5 paginePsicologia MusicaleAttilio PerfettiNessuna valutazione finora
- Grandi Vie NervoseDocumento15 pagineGrandi Vie NervoseAnonymous 9aq7eNvONessuna valutazione finora
- Curs MCurs Mario Castagnini - Educare Neuromotorieario Castagnini - Educare NeuromotorieDocumento138 pagineCurs MCurs Mario Castagnini - Educare Neuromotorieario Castagnini - Educare NeuromotorieBiborka GyepesiNessuna valutazione finora
- Midollo TroncoDocumento15 pagineMidollo Troncoeve_612Nessuna valutazione finora
- NEUROANATOMIADocumento172 pagineNEUROANATOMIAFrancesco LanzaNessuna valutazione finora