Potrebbero piacerti anche
- Prontuario di lotta biologica contro i nemici dell’orto: Combattere e sconfiggere i parassiti che aggrediscono i nostri ortiDa EverandProntuario di lotta biologica contro i nemici dell’orto: Combattere e sconfiggere i parassiti che aggrediscono i nostri ortiNessuna valutazione finora
- Funghiamo? L’arte del cercare e raccoglier funghiDa EverandFunghiamo? L’arte del cercare e raccoglier funghiNessuna valutazione finora
- Lillo La Chiusa - Guida Completa Al Riconoscimento Dei Funghi-De Vecchi (2020)Documento351 pagineLillo La Chiusa - Guida Completa Al Riconoscimento Dei Funghi-De Vecchi (2020)santbasNessuna valutazione finora
- Ottenimento Ibridi InterspecificiDocumento90 pagineOttenimento Ibridi InterspecificiOrtensio LuongoNessuna valutazione finora
- Le Caratteristiche Dei FunghiDocumento8 pagineLe Caratteristiche Dei FunghiRaffaello PrandoNessuna valutazione finora
- Copia Di Piante BioindicatriciDocumento43 pagineCopia Di Piante BioindicatriciMaurizio Maurizio Kart0% (1)
- BOTANICADocumento20 pagineBOTANICAGiuseppe GriecoNessuna valutazione finora
- UD1 4 FunghiDocumento21 pagineUD1 4 FunghiDavide SalvestroniNessuna valutazione finora
- 4 5-AfidiDocumento12 pagine4 5-AfidiGerardo SuglianoNessuna valutazione finora
- 01 ProtozoiDocumento19 pagine01 ProtozoiEugenio StracuzzaNessuna valutazione finora
- I DeuteromicetiDocumento16 pagineI DeuteromicetiJacopo ParavanNessuna valutazione finora
- Conoscere I Funghi PDFDocumento164 pagineConoscere I Funghi PDFIvan MontellaNessuna valutazione finora
- ErbarioDocumento39 pagineErbarioMistyDustNessuna valutazione finora
- La Classificazione Delle PianteDocumento4 pagineLa Classificazione Delle PianteTiziana ScuderiNessuna valutazione finora
- Botanica DispenseDocumento13 pagineBotanica DispenseGiorgioNessuna valutazione finora
- Alghe Azzurre (P)Documento66 pagineAlghe Azzurre (P)tizlombNessuna valutazione finora
- Riassunti MICOLOGIADocumento22 pagineRiassunti MICOLOGIAValeria Di StefanoNessuna valutazione finora
- Anticrittogamici Detti Fungicidi Definitivo Stop!Documento52 pagineAnticrittogamici Detti Fungicidi Definitivo Stop!nat copNessuna valutazione finora
- Botanica 2Documento2 pagineBotanica 2Eleonora CrespiaticoNessuna valutazione finora
- AscomicetiDocumento8 pagineAscomicetiGiorgioNessuna valutazione finora
- MicologiaDocumento13 pagineMicologiaMassimiliano PalumboNessuna valutazione finora
- I ProtistiDocumento15 pagineI ProtistiFederico GinocchiNessuna valutazione finora
- Botanica BriofiteDocumento3 pagineBotanica BriofiteAdriana PellitteriNessuna valutazione finora
- Le Piante-1Documento3 pagineLe Piante-1salmaNessuna valutazione finora
- AGC Guida Preparazione Esame Abilitazione Ricerca e Raccolta Tartufi PDFDocumento61 pagineAGC Guida Preparazione Esame Abilitazione Ricerca e Raccolta Tartufi PDFStefano PapittoNessuna valutazione finora
- 35 Batteri, Protisti e FunghiDocumento5 pagine35 Batteri, Protisti e FunghiCrocetti SimonaNessuna valutazione finora
- AFIDIDocumento3 pagineAFIDIpierluighinaNessuna valutazione finora
- Riconoscere e Combattere Le Principali Malattie Fungine Delle PianteDocumento8 pagineRiconoscere e Combattere Le Principali Malattie Fungine Delle PianteLuciano Gianfranco PaianolNessuna valutazione finora
- Storia Delle OrchideeDocumento13 pagineStoria Delle Orchideenagual973Nessuna valutazione finora
- Crittogame VascolariDocumento28 pagineCrittogame VascolariAnna De Simone100% (2)
- 1 Il Regno Dei FunghiDocumento2 pagine1 Il Regno Dei FunghiChiara L.Nessuna valutazione finora
- AngiospermeDocumento12 pagineAngiospermeDaniele De PietriNessuna valutazione finora
- Botanica IDocumento42 pagineBotanica ISilvia DadoNessuna valutazione finora
- Dispensa Di BotanicaDocumento8 pagineDispensa Di BotanicaStefano DestefaniNessuna valutazione finora
- Melo 01 01Documento21 pagineMelo 01 01TransmontesoNessuna valutazione finora
- DEUTEROMICETIDocumento10 pagineDEUTEROMICETIGiorgioNessuna valutazione finora
- Erboristeria Erbario Fitopatologico PDFDocumento39 pagineErboristeria Erbario Fitopatologico PDFswamy88Nessuna valutazione finora
- Produzioni Vegetali Parte 2, Appunti Scienze Dell'alimentazioneDocumento21 pagineProduzioni Vegetali Parte 2, Appunti Scienze Dell'alimentazioneGiorgio SciarraNessuna valutazione finora
- BASIDIOMICETIDocumento5 pagineBASIDIOMICETIGiorgioNessuna valutazione finora
- Il Regno Dei FunghiDocumento16 pagineIl Regno Dei Funghipipo piposoNessuna valutazione finora
- Frutti Secchi DeiscentiDocumento86 pagineFrutti Secchi DeiscentiAmanda HartNessuna valutazione finora
- GLOSSARIO BotanicaDocumento24 pagineGLOSSARIO BotanicaMatteo PeruzzoNessuna valutazione finora
- FloraDocumento7 pagineFloraENKIINessuna valutazione finora
- 13 I MicetiDocumento4 pagine13 I MicetifabiothekingjamesNessuna valutazione finora
- Dispensa Corso MicologicoDocumento51 pagineDispensa Corso MicologicoWalter LentiniNessuna valutazione finora
- Classificazione Degli Esseri ViventiDocumento72 pagineClassificazione Degli Esseri Viventiariel cinturaNessuna valutazione finora
- Micologia+Batteriologia SpecialeDocumento56 pagineMicologia+Batteriologia SpecialeFilippo BiondiNessuna valutazione finora
- AnetoDocumento9 pagineAnetoacidexp66Nessuna valutazione finora
- E BOOK Legambiente-FestadellAlbero - 2020Documento24 pagineE BOOK Legambiente-FestadellAlbero - 2020Legambiente SalveNessuna valutazione finora
- Alpicoltura - Dispensa GusmeroliDocumento92 pagineAlpicoltura - Dispensa GusmeroliElia LipreriNessuna valutazione finora
- GimnospermeDocumento45 pagineGimnospermeChiara Cubbins GambardellaNessuna valutazione finora
- Piante Grasse - GiuntiDocumento130 paginePiante Grasse - Giuntimartina palumboNessuna valutazione finora
- ZIGOMICETI e DERMATOFITIDocumento10 pagineZIGOMICETI e DERMATOFITILiv TartaglioneNessuna valutazione finora
- Lezione 18Documento15 pagineLezione 18Paolo CNessuna valutazione finora
- 03-Briofite SC AmbDocumento77 pagine03-Briofite SC AmbMarco De LucaNessuna valutazione finora
- Mediterranean Proper Flora Critica D Italia HydrophyllaceaeDocumento23 pagineMediterranean Proper Flora Critica D Italia HydrophyllaceaeAnte BuraNessuna valutazione finora
- Conoscere I FunghiDocumento164 pagineConoscere I FunghiemilioNessuna valutazione finora
- CicoriaDocumento4 pagineCicoriawilliamNessuna valutazione finora
- I funghi mangerecci e velenosi dell'Europa media con speciale riguardo a quelli che crescono nel Trentino - II edizione riveduta ed aumentataDa EverandI funghi mangerecci e velenosi dell'Europa media con speciale riguardo a quelli che crescono nel Trentino - II edizione riveduta ed aumentataNessuna valutazione finora
- Yogurt BNDocumento10 pagineYogurt BNabyutzaNessuna valutazione finora
- Tehnologii Ecologice de Cultivare A LegumelorDocumento43 pagineTehnologii Ecologice de Cultivare A LegumelorabyutzaNessuna valutazione finora
- 1 Industrie AgrarieDocumento36 pagine1 Industrie AgrarieabyutzaNessuna valutazione finora
- 01 Introd Uzi OneDocumento53 pagine01 Introd Uzi OneabyutzaNessuna valutazione finora
- 8 Lattosio BNDocumento18 pagine8 Lattosio BNabyutzaNessuna valutazione finora
- Genetica APIDocumento21 pagineGenetica APIabyutzaNessuna valutazione finora
- 2 Pagam Latte QualitaDocumento13 pagine2 Pagam Latte QualitaabyutzaNessuna valutazione finora
- 9-Latte Alimentare BNDocumento21 pagine9-Latte Alimentare BNabyutzaNessuna valutazione finora
- Genetica Agraria 2 (Modalità Compatibilità)Documento77 pagineGenetica Agraria 2 (Modalità Compatibilità)abyutzaNessuna valutazione finora
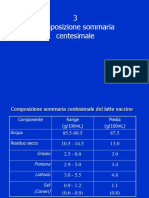
- 3 Composiz - Sommaria Instab FasiDocumento14 pagine3 Composiz - Sommaria Instab FasiabyutzaNessuna valutazione finora
- AtlanteIrrigazione NETDocumento178 pagineAtlanteIrrigazione NETabyutzaNessuna valutazione finora
- Lez 10 - Virus PDFDocumento52 pagineLez 10 - Virus PDFabyutzaNessuna valutazione finora
- AtlanteIrrigazione NETDocumento178 pagineAtlanteIrrigazione NETabyutzaNessuna valutazione finora
- Genetica Di Popolazione (Modalità Compatibilità)Documento27 pagineGenetica Di Popolazione (Modalità Compatibilità)abyutzaNessuna valutazione finora
- Classica2 (Modalità Compatibilità)Documento11 pagineClassica2 (Modalità Compatibilità)abyutzaNessuna valutazione finora
- 2 RichiamiMeccFluidiDocumento28 pagine2 RichiamiMeccFluidiabyutzaNessuna valutazione finora
- Pluv RossanoDocumento1 paginaPluv RossanoabyutzaNessuna valutazione finora
- 1 SistemiIrriguiDocumento69 pagine1 SistemiIrriguiabyutzaNessuna valutazione finora
- 4 IdrodinamicaDocumento74 pagine4 IdrodinamicaabyutzaNessuna valutazione finora
- 5 Liquidi RealiDocumento25 pagine5 Liquidi RealiabyutzaNessuna valutazione finora
- Il Brusone Del RisoDocumento13 pagineIl Brusone Del RisoabyutzaNessuna valutazione finora
- Lez 6 - Germinazione Spore e Ciclo Di PatogenesiDocumento19 pagineLez 6 - Germinazione Spore e Ciclo Di PatogenesiabyutzaNessuna valutazione finora
- 3 IdrostaticaDocumento39 pagine3 IdrostaticaabyutzaNessuna valutazione finora
- 6 CorrentiPressioneDocumento54 pagine6 CorrentiPressioneabyutzaNessuna valutazione finora
- Lez 3 - SintomatologiaDocumento30 pagineLez 3 - SintomatologiaabyutzaNessuna valutazione finora
- Rassegna Cancro CastagnoDocumento22 pagineRassegna Cancro CastagnoabyutzaNessuna valutazione finora
- Lez 18 Resistenza Ai FungicidiDocumento18 pagineLez 18 Resistenza Ai FungicidiabyutzaNessuna valutazione finora
- Lez 10 - Virus PDFDocumento52 pagineLez 10 - Virus PDFabyutzaNessuna valutazione finora
- L'Oidio Della Vite in ToscanaDocumento31 pagineL'Oidio Della Vite in ToscanaabyutzaNessuna valutazione finora
- CUORE MATTO-Little TonyDocumento3 pagineCUORE MATTO-Little TonyluxNessuna valutazione finora
- Un Milioneottocentomila Passi. Io, Il Mio Bambino e Il Cammino Di SantiagoDocumento2 pagineUn Milioneottocentomila Passi. Io, Il Mio Bambino e Il Cammino Di SantiagoElisabetta OrlandiNessuna valutazione finora
- PoesieDocumento58 paginePoesiealfredo89Nessuna valutazione finora
- Fotografia Digitale. Io Parto Da ZeroDocumento150 pagineFotografia Digitale. Io Parto Da ZeroCristian MancusoNessuna valutazione finora