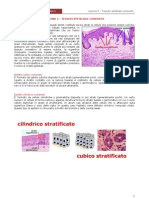
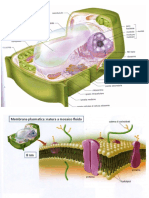
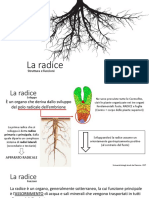
Potrebbero piacerti anche
- Appunti Di Biologia Vegetale. Istologia e Anatomia VegetaleDocumento120 pagineAppunti Di Biologia Vegetale. Istologia e Anatomia VegetaleCOM srl100% (2)
- Corso Potatura DispensaDocumento105 pagineCorso Potatura Dispensacharles radaoniNessuna valutazione finora
- Tre Trattati Di AlchimiaDocumento29 pagineTre Trattati Di AlchimiaclekupreNessuna valutazione finora
- Jules Renard - Pel Di Carota (1894)Documento123 pagineJules Renard - Pel Di Carota (1894)nastel100% (1)
- 4 Tracheofite PteridofiteDocumento8 pagine4 Tracheofite PteridofiteAdriano SpiritoNessuna valutazione finora
- Controlul Calitatii SmantaniiDocumento33 pagineControlul Calitatii SmantaniiPop Mihai100% (2)
- I Tessuti VegetaliDocumento13 pagineI Tessuti Vegetalialice9980Nessuna valutazione finora
- Tessuti AdultiDocumento3 pagineTessuti AdultiRosanna Di CarloNessuna valutazione finora
- Botanica GeneraleDocumento103 pagineBotanica GeneraleChiara PapagnoNessuna valutazione finora
- Tessuti MeristematiciDocumento1 paginaTessuti MeristematiciChristian BertoglioNessuna valutazione finora
- 4 Il FustoDocumento4 pagine4 Il FustoAsia ZaccariaNessuna valutazione finora
- 3 - Anatomia-ComparataDocumento14 pagine3 - Anatomia-ComparataEugenio MolestiNessuna valutazione finora
- Anatomia VegetaleDocumento8 pagineAnatomia VegetaleChiara PellitteriNessuna valutazione finora
- Bahan Persentasi BiologoDocumento17 pagineBahan Persentasi BiologoSilva RahmaNessuna valutazione finora
- Classificazione Dei Regni:: AutotofriDocumento20 pagineClassificazione Dei Regni:: AutotofriNicolò StramondoNessuna valutazione finora
- Botanica e Biologia GeneraleDocumento23 pagineBotanica e Biologia GeneraleCarloTheShowmanRopertoNessuna valutazione finora
- 2 I TessutiDocumento3 pagine2 I TessutiAsia ZaccariaNessuna valutazione finora
- Atlante BotanicaDocumento13 pagineAtlante BotanicaMartina TresoldiNessuna valutazione finora
- Botanica BriofiteDocumento3 pagineBotanica BriofiteAdriana PellitteriNessuna valutazione finora
- 1 La Cellula VegetaleDocumento3 pagine1 La Cellula VegetaleAsia ZaccariaNessuna valutazione finora
- 08 Organi VegetaliDocumento34 pagine08 Organi VegetaliPacifico85 el magicoNessuna valutazione finora
- Aronne 7 Apici Vegetativi RadicaliDocumento1 paginaAronne 7 Apici Vegetativi RadicalibvincenzoNessuna valutazione finora
- BotanicaDocumento40 pagineBotanicaArturo FiorenzaNessuna valutazione finora
- Tessuto TegumentaleDocumento26 pagineTessuto TegumentaleSlavica Džerfi MatanovicNessuna valutazione finora
- Biologia Vegetale Tomo 2Documento35 pagineBiologia Vegetale Tomo 2Antonio DanieleNessuna valutazione finora
- Ve TriniDocumento10 pagineVe TriniMartina TresoldiNessuna valutazione finora
- PIANTEDocumento7 paginePIANTEValeria Di StefanoNessuna valutazione finora
- BotanicaDocumento6 pagineBotanicaPaolo BrignoliNessuna valutazione finora
- L'anatomia Delle PianteDocumento17 pagineL'anatomia Delle PianteIrene MazzolaNessuna valutazione finora
- BotanicaDocumento8 pagineBotanicaChiara IannielloNessuna valutazione finora
- Lezione 2 - Tessuto Epiteliale CompostoDocumento7 pagineLezione 2 - Tessuto Epiteliale CompostotrillicosaNessuna valutazione finora
- Tessuti Tegumentali IIDocumento7 pagineTessuti Tegumentali IIAnna De SimoneNessuna valutazione finora
- Terza LezioneDocumento6 pagineTerza LezioneFederica MaiolicaNessuna valutazione finora
- Tessuti Tegumentali IDocumento4 pagineTessuti Tegumentali IAnna De SimoneNessuna valutazione finora
- BOTANICADocumento20 pagineBOTANICAGiuseppe GriecoNessuna valutazione finora
- LA CellulaDocumento3 pagineLA CellulaCosso GiuliaNessuna valutazione finora
- Botanica 2Documento2 pagineBotanica 2Eleonora CrespiaticoNessuna valutazione finora
- SpermatofiteDocumento4 pagineSpermatofiteAdriana PellitteriNessuna valutazione finora
- Anatomia FustoDocumento6 pagineAnatomia FustoEnrica EspositoNessuna valutazione finora
- Crescita e Forma Degli AlberiDocumento9 pagineCrescita e Forma Degli AlberiAlfraedNessuna valutazione finora
- Biologia VegataleDocumento9 pagineBiologia VegataleiwttbaNessuna valutazione finora
- 5.CAPITOLO 22-GimnospermeDocumento6 pagine5.CAPITOLO 22-GimnospermeBeatrice EvangelistiNessuna valutazione finora
- Anatomia - Anatomia MicroscopicaDocumento38 pagineAnatomia - Anatomia MicroscopicaiccitinderNessuna valutazione finora
- 1- parenchimi_02-4-19Documento61 pagine1- parenchimi_02-4-19tomaoana011Nessuna valutazione finora
- La Cellula VegetaleDocumento47 pagineLa Cellula VegetaleEleonora AngelocolaNessuna valutazione finora
- La RadiceDocumento45 pagineLa RadiceTiziana ToniniNessuna valutazione finora
- Le Piante AngiospermeDocumento5 pagineLe Piante AngiospermeAnouk Peña SvaldiNessuna valutazione finora
- Appunti IstologiaDocumento28 pagineAppunti IstologiaJennifer SghedoniNessuna valutazione finora
- Copia Di Piante BioindicatriciDocumento43 pagineCopia Di Piante BioindicatriciMaurizio Maurizio Kart0% (1)
- BiologiaDocumento61 pagineBiologianuny91Nessuna valutazione finora
- Sostanze Vegetali Farmaceutiche UnicoDocumento139 pagineSostanze Vegetali Farmaceutiche Unico20047230Nessuna valutazione finora
- ISTOLOGIADocumento9 pagineISTOLOGIAAndrea MancaNessuna valutazione finora
- Piante Con CruciverbaDocumento5 paginePiante Con CruciverbaRoberto StampettaNessuna valutazione finora
- Radici, Fusto e Foglie - Wednesday, Jan. 15 - InstapaperDocumento14 pagineRadici, Fusto e Foglie - Wednesday, Jan. 15 - InstapaperpatriziapegoloNessuna valutazione finora
- Crittogame VascolariDocumento28 pagineCrittogame VascolariAnna De Simone100% (2)
- IstologiaDocumento26 pagineIstologiaFederica RiiliNessuna valutazione finora
- Scienze Corpo UmanoDocumento7 pagineScienze Corpo UmanoGiulia MaestriniNessuna valutazione finora
- 08 ArtropodiDocumento10 pagine08 ArtropodiEugenio StracuzzaNessuna valutazione finora
- 06 Tessuti EpitelialiDocumento6 pagine06 Tessuti EpitelialifradoctorNessuna valutazione finora
- Le Cellule Eucariotiche - BookinprogressDocumento12 pagineLe Cellule Eucariotiche - BookinprogressCarmenNessuna valutazione finora
- Botanica BatteriDocumento4 pagineBotanica BatteriAdriana PellitteriNessuna valutazione finora
- 2b La Foglia 2Documento6 pagine2b La Foglia 2HajarNessuna valutazione finora
- Leonardo Sciascia - Atti Relativi Alla Morte Di Raymond Roussel (Ita Libro)Documento16 pagineLeonardo Sciascia - Atti Relativi Alla Morte Di Raymond Roussel (Ita Libro)davanto0Nessuna valutazione finora
- Diritto Dell'ambiente - N.LugaresiDocumento20 pagineDiritto Dell'ambiente - N.Lugaresiget100% (1)
- Mappa Concettuale. DurkheimDocumento12 pagineMappa Concettuale. DurkheimIulianButnaruNessuna valutazione finora
- Massimiliano Gaetano - Le Qualità Dell'animo in Astrologia - Luna e Mercurio PDFDocumento10 pagineMassimiliano Gaetano - Le Qualità Dell'animo in Astrologia - Luna e Mercurio PDFMassimiliano GaetanoNessuna valutazione finora
- Tutti Gli Uomini Che Hanno Fatto La Cultura Di DestraDocumento2 pagineTutti Gli Uomini Che Hanno Fatto La Cultura Di DestraAlberto SamonàNessuna valutazione finora
- Tesi WipDocumento44 pagineTesi WipGiuseppe Tyler CinquegranaNessuna valutazione finora
- Il Sistema Di Credenze e I Quattro (Cinque Accordi)Documento4 pagineIl Sistema Di Credenze e I Quattro (Cinque Accordi)Laura RobinsonNessuna valutazione finora
- 12 Mesi - BRESCIA - Ottobre 2012Documento99 pagine12 Mesi - BRESCIA - Ottobre 201212mesiNessuna valutazione finora
- B2 - La Voce Del Padrone - BattiatoDocumento5 pagineB2 - La Voce Del Padrone - BattiatoEmanuele MastrangeliNessuna valutazione finora
- 02 Antropologia e NazionalismoDocumento10 pagine02 Antropologia e Nazionalismoapi-3735634100% (1)
- CHN3 - Regolamento Tunnel & Troll in Italiano - Ed. Chimerae Hobby Group - Tunnels & Trolls T&TDocumento8 pagineCHN3 - Regolamento Tunnel & Troll in Italiano - Ed. Chimerae Hobby Group - Tunnels & Trolls T&Tjasc0_hotmail_itNessuna valutazione finora
- 26 - Anatomia II - 30-03-2017 (R)Documento10 pagine26 - Anatomia II - 30-03-2017 (R)SandroNessuna valutazione finora