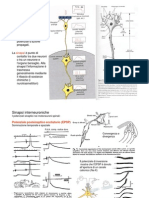
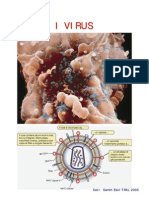
Potrebbero piacerti anche
- 10 Silenziamento GenicoDocumento21 pagine10 Silenziamento GenicoGloria Ashes NapoliNessuna valutazione finora
- Lezione XIV. Analisi Molecolare Dei GeniDocumento38 pagineLezione XIV. Analisi Molecolare Dei GenigiudisNessuna valutazione finora
- Scienze Dal Dna Alle ProteineDocumento10 pagineScienze Dal Dna Alle ProteineNo NiNessuna valutazione finora
- Genetica MedicaDocumento151 pagineGenetica MedicaBeatrice CarliniNessuna valutazione finora
- Sintesi ProteicaDocumento8 pagineSintesi ProteicamicheleNessuna valutazione finora
- Isolamento GenicoDocumento19 pagineIsolamento GenicoRobNessuna valutazione finora
- UntitledDocumento2 pagineUntitledRosi De LucaNessuna valutazione finora
- Lez 9 - Il NucleoDocumento46 pagineLez 9 - Il NucleosaretttNessuna valutazione finora
- Sintesi ProteineDocumento53 pagineSintesi Proteinesimeone.salvatoreleonardoNessuna valutazione finora
- F00a-Individualità GeneticaDocumento16 pagineF00a-Individualità Geneticaantonio nennaNessuna valutazione finora
- Il GenomaDocumento2 pagineIl GenomaSimona CafàNessuna valutazione finora
- Librerie GenomicheDocumento1 paginaLibrerie GenomicheGiordana GiampaoloNessuna valutazione finora
- Bio AnimaleDocumento29 pagineBio AnimaleGiulia DusettiNessuna valutazione finora
- C 3 P 45 Sintesi ProteicaDocumento2 pagineC 3 P 45 Sintesi Proteicagiperico7378Nessuna valutazione finora
- 14-L22Paronetto Caporossi Trascrizione e TraduzioneDocumento71 pagine14-L22Paronetto Caporossi Trascrizione e TraduzioneCarlosNessuna valutazione finora
- Il Metodo Crispr Cas9Documento15 pagineIl Metodo Crispr Cas9Federica PicchiNessuna valutazione finora
- 2015 Lezione 5 - ReplicazioneDocumento69 pagine2015 Lezione 5 - Replicazionemari mateiNessuna valutazione finora
- La Genomica ComparativaDocumento5 pagineLa Genomica ComparativaAnna CrestaniNessuna valutazione finora
- Trasmissione MendelianaDocumento129 pagineTrasmissione MendelianaVito DamascoNessuna valutazione finora
- Codice Genetico e Traduzione PDFDocumento58 pagineCodice Genetico e Traduzione PDFGaia Mitsuko PrioloNessuna valutazione finora
- 6) Sintesi Delle ProteineDocumento1 pagina6) Sintesi Delle Proteineinksuky.3dNessuna valutazione finora
- Regolazione TrascrizionaleDocumento65 pagineRegolazione TrascrizionaleAlessandro PiccottiNessuna valutazione finora
- Virus A RNA1Documento21 pagineVirus A RNA1msjackellNessuna valutazione finora
- Biologia Capitolo 4Documento55 pagineBiologia Capitolo 4Arianna ManderioliNessuna valutazione finora
- Dna ContinuoDocumento11 pagineDna ContinuoPamelaNessuna valutazione finora
- Dna E Rna: Eleonora LacquanitiDocumento10 pagineDna E Rna: Eleonora Lacquanitieleonora.lacquaniti.123Nessuna valutazione finora
- Capitolo 10Documento1 paginaCapitolo 10Nic15Z11Nessuna valutazione finora
- Lezione 3Documento72 pagineLezione 3DS87Nessuna valutazione finora
- 1 - Dna PDFDocumento78 pagine1 - Dna PDFMariagiovanna MormileNessuna valutazione finora
- Battaglioli BiologiaCellulare Lez3Documento67 pagineBattaglioli BiologiaCellulare Lez3Renata WürziusNessuna valutazione finora
- Splicing AlternativoDocumento52 pagineSplicing AlternativoRCNessuna valutazione finora
- Lezione 7 - RNA e TrascrizioneDocumento11 pagineLezione 7 - RNA e TrascrizioneMartina BaroliNessuna valutazione finora
- 05 181016.pptx LockedDocumento14 pagine05 181016.pptx Lockedtemaj95108Nessuna valutazione finora
- 09 261016.pptx LockedDocumento14 pagine09 261016.pptx Lockedtemaj95108Nessuna valutazione finora
- Paniere Domande Multiple Biologia ApplicataDocumento24 paginePaniere Domande Multiple Biologia ApplicatamartinaNessuna valutazione finora
- Dna e CromosomiDocumento3 pagineDna e Cromosomigiulia maria raspaNessuna valutazione finora
- Replicazione Di Un Gene l5Documento8 pagineReplicazione Di Un Gene l5aldo jacoNessuna valutazione finora
- Duplicazione DNA e Sintesi Proteica 2Documento22 pagineDuplicazione DNA e Sintesi Proteica 2Giulia CaliriNessuna valutazione finora
- Lezione 26Documento6 pagineLezione 26Mariacarmela MorrealeNessuna valutazione finora
- MirnaDocumento15 pagineMirnaTeresa CicconeNessuna valutazione finora
- Domande Frequenti BiologiaDocumento3 pagineDomande Frequenti Biologiaboh2345Nessuna valutazione finora
- BiologiaDocumento8 pagineBiologiala raNessuna valutazione finora
- BIOTECNOLOGIEDocumento6 pagineBIOTECNOLOGIEfrancesca sicaliNessuna valutazione finora
- La Sintesi Proteica: Un Lavoro Di Precisione: La Vita e La CellulaDocumento2 pagineLa Sintesi Proteica: Un Lavoro Di Precisione: La Vita e La CellulaFedericaNessuna valutazione finora
- Genom I GenDocumento17 pagineGenom I GenGoran100% (1)
- Gen pt1Documento19 pagineGen pt1marietto002Nessuna valutazione finora
- Codice Genetico-Traduzione-Sintesi ProteicaDocumento39 pagineCodice Genetico-Traduzione-Sintesi Proteicamolesdan3540Nessuna valutazione finora
- L'ereditarietà e Il GenomaDocumento12 pagineL'ereditarietà e Il GenomaLeunora HoxhaNessuna valutazione finora
- DNA e RNADocumento12 pagineDNA e RNALuca BaroniNessuna valutazione finora
- Lez 3 MicroDocumento12 pagineLez 3 Microclacelove2004Nessuna valutazione finora
- Retrovirus e HIVDocumento96 pagineRetrovirus e HIVpattada3333Nessuna valutazione finora
- Biologia Molecolare 1 07-08Documento26 pagineBiologia Molecolare 1 07-08Loni RodriguezNessuna valutazione finora
- Capitoli B3 e B4Documento11 pagineCapitoli B3 e B4Gabriele PalermoNessuna valutazione finora
- Lezione 10Documento5 pagineLezione 10yazan odehNessuna valutazione finora
- Esame Bio Mol 04-06 Prima CorrezioneDocumento6 pagineEsame Bio Mol 04-06 Prima CorrezioneAu VeNessuna valutazione finora
- Concetti Di Genetica MedicaDocumento28 pagineConcetti Di Genetica Medicajuri84Nessuna valutazione finora
- 8 Vettori e ClonaggioDocumento6 pagine8 Vettori e ClonaggiobvincenzoNessuna valutazione finora
- Oltre il DNA: La Rivoluzione Epigenetica: Da Meccanismi Cellulari a Fattori Ambientali: Come l'Epigenetica Plasmi il Nostro Destino Biologico e le Implicazioni per la Salute, il Comportamento e il Futuro della RicercaDa EverandOltre il DNA: La Rivoluzione Epigenetica: Da Meccanismi Cellulari a Fattori Ambientali: Come l'Epigenetica Plasmi il Nostro Destino Biologico e le Implicazioni per la Salute, il Comportamento e il Futuro della RicercaNessuna valutazione finora
- Trasmissione SinapticaDocumento12 pagineTrasmissione Sinapticaapi-3774792100% (1)
- Contrazione MuscolareDocumento15 pagineContrazione Muscolareapi-26155364100% (4)
- 9 - Recettori Sensoriali, Sistemi IntegratiDocumento14 pagine9 - Recettori Sensoriali, Sistemi Integratiapi-26155364Nessuna valutazione finora
- 7 - Neuroni, Integrazione, NeuromodulazioneDocumento12 pagine7 - Neuroni, Integrazione, Neuromodulazioneapi-26155364Nessuna valutazione finora
- ElettrofisiologiaDocumento12 pagineElettrofisiologiaapi-3774792100% (1)
- 3 - Trasporto AttivoDocumento9 pagine3 - Trasporto Attivoapi-26155364Nessuna valutazione finora
- Fisiologia Parte 4.pdaDocumento14 pagineFisiologia Parte 4.pdaapi-3774792Nessuna valutazione finora
- TUBERCOLOSI Lanciotti 08 OKDocumento8 pagineTUBERCOLOSI Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- RischioBiol LoNostro 06Documento62 pagineRischioBiol LoNostro 06api-26155364Nessuna valutazione finora
- 1 - Introduzione e MembranaDocumento28 pagine1 - Introduzione e Membranaapi-26155364Nessuna valutazione finora
- Domande Frequenti PazzagliDocumento1 paginaDomande Frequenti Pazzagliapi-26155364Nessuna valutazione finora
- Lab Biochimica PaoliDocumento28 pagineLab Biochimica Paoliapi-26155364Nessuna valutazione finora
- 2 - Eq - ElettrochimDocumento6 pagine2 - Eq - Elettrochimapi-26155364Nessuna valutazione finora
- Virus 08 PPTDocumento26 pagineVirus 08 PPTapi-26155364Nessuna valutazione finora
- ProgrammaDocumento2 pagineProgrammaapi-26155364Nessuna valutazione finora
- TBC Iss 06Documento4 pagineTBC Iss 06api-26155364Nessuna valutazione finora
- Tifo e Paratifo - Lanciotti08Documento8 pagineTifo e Paratifo - Lanciotti08api-26155364Nessuna valutazione finora
- SPORIGENI&TETANO Lanciotti 08 OKDocumento6 pagineSPORIGENI&TETANO Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- TBC BonanniDocumento30 pagineTBC Bonanniapi-26155364Nessuna valutazione finora
- Poliomielite CrovariDocumento32 paginePoliomielite Crovariapi-26155364Nessuna valutazione finora
- RischioInf Viaggiatori 08Documento20 pagineRischioInf Viaggiatori 08api-26155364Nessuna valutazione finora
- TBC BonanniDocumento30 pagineTBC Bonanniapi-26155364100% (2)
- PRIONI&BSE Lanciotti 08 OKDocumento9 paginePRIONI&BSE Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- HB InfermieriDocumento22 pagineHB Infermieriapi-26155364100% (1)
- HB InfermieriDocumento22 pagineHB Infermieriapi-26155364100% (1)
- INFLUENZA Lanciotti 08 OKDocumento47 pagineINFLUENZA Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- Meningiti ISS 06Documento2 pagineMeningiti ISS 06api-26155364Nessuna valutazione finora
- Influenza CrovariDocumento48 pagineInfluenza Crovariapi-26155364Nessuna valutazione finora
- INFLUENZA Lanciotti 08 OKDocumento47 pagineINFLUENZA Lanciotti 08 OKapi-26155364Nessuna valutazione finora
- EpatitiCroniche 06 OKDocumento23 pagineEpatitiCroniche 06 OKapi-26155364Nessuna valutazione finora