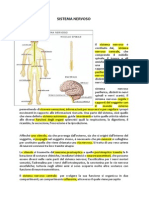
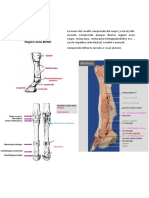
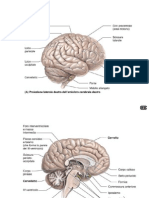
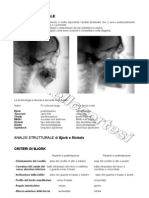
Potrebbero piacerti anche
- Riassunto Bear Neuroscienze Esplorando Il CervelloDocumento9 pagineRiassunto Bear Neuroscienze Esplorando Il CervellocdeipoliNessuna valutazione finora
- Neuroanatomia - Riassunto Del CattaneoDocumento47 pagineNeuroanatomia - Riassunto Del CattaneoAlessandro Fazio50% (2)
- PROTOCOLLO FRENULO Marchesan Anna BeghettoDocumento12 paginePROTOCOLLO FRENULO Marchesan Anna BeghettoAnonymous 1V3LWvE100% (1)
- Anatomia 2Documento58 pagineAnatomia 2GioNessuna valutazione finora
- Fisiologia Del Parto Completo Colacurci 5 OttobreDocumento49 pagineFisiologia Del Parto Completo Colacurci 5 OttobreLina Coppola0% (1)
- Lezioni Vie Nervose SNAutonomoDocumento80 pagineLezioni Vie Nervose SNAutonomoVincenzo BinantiNessuna valutazione finora
- Fisiopatologia Sistema Nervoso Autonomo PDFDocumento10 pagineFisiopatologia Sistema Nervoso Autonomo PDFpaolo68audisioNessuna valutazione finora
- Vie SensitiveDocumento21 pagineVie SensitivestefpecNessuna valutazione finora
- Neuroanatomia RiassuntoDocumento5 pagineNeuroanatomia RiassuntoAlice1102Nessuna valutazione finora
- Funzione MotoriaDocumento19 pagineFunzione MotoriaEdoardo BerteroNessuna valutazione finora
- Vie SomatosensorialiDocumento4 pagineVie Somatosensorialidimilda100% (1)
- Vie NervoseDocumento5 pagineVie Nervosexxx9090Nessuna valutazione finora
- Sistema Nervoso Periferico - Nervi CraniciDocumento10 pagineSistema Nervoso Periferico - Nervi CraniciSabrina CillariNessuna valutazione finora
- Sostanza GrigiaDocumento4 pagineSostanza GrigiaAnthony NicosiaNessuna valutazione finora
- 7 Vie Senso 2018Documento90 pagine7 Vie Senso 2018Chloe BujuoirNessuna valutazione finora
- Riassunti Sistema NervosoDocumento40 pagineRiassunti Sistema NervosoMarisa TalaricoNessuna valutazione finora
- FISIOLOGIADocumento6 pagineFISIOLOGIAMariateresa SantorsolaNessuna valutazione finora
- Scienze BiomedicheDocumento21 pagineScienze BiomedicheRenato RossiNessuna valutazione finora
- Sistema Nervoso (RIASSUNTO)Documento6 pagineSistema Nervoso (RIASSUNTO)Dani Lo MagroNessuna valutazione finora
- Neurologia PecorelliDocumento141 pagineNeurologia Pecorellibasturk81Nessuna valutazione finora
- Domande NeuroanatomiaDocumento3 pagineDomande NeuroanatomiaHermannNessuna valutazione finora
- Sistema Nervoso RiassuntoDocumento64 pagineSistema Nervoso Riassuntolucchini.paolo3002Nessuna valutazione finora
- Schemi Sintetici Delle Vie NervoseDocumento17 pagineSchemi Sintetici Delle Vie NervoseSalvo Danilo LombardoNessuna valutazione finora
- Anatomia e Fisopatologia 2021Documento137 pagineAnatomia e Fisopatologia 2021Ejona SabijaNessuna valutazione finora
- Grandi Vie NervoseDocumento15 pagineGrandi Vie NervoseAnonymous 9aq7eNvONessuna valutazione finora
- Dispensa Acustica CervelloDocumento20 pagineDispensa Acustica CervelloAlfonso BelfioreNessuna valutazione finora
- Il Midollo Spinale - Amyotrophic Lateral Sclerosis PDFDocumento6 pagineIl Midollo Spinale - Amyotrophic Lateral Sclerosis PDFAngelo Dell'AnnaNessuna valutazione finora
- Motorni Putevi Cns-ADocumento8 pagineMotorni Putevi Cns-ABožidar Ćaćić100% (1)
- Tecniche Della Riabilitazione GraficaDocumento27 pagineTecniche Della Riabilitazione Graficagiorgia.sacchieroNessuna valutazione finora
- Plesso BrachialeDocumento9 paginePlesso BrachialeantonioNessuna valutazione finora
- Sistema NervosoDocumento14 pagineSistema NervosoMiurrinaNessuna valutazione finora
- Sistema SensorimotorioDocumento6 pagineSistema SensorimotorioMiriam LongobardiNessuna valutazione finora
- Nervi CraniciDocumento31 pagineNervi CraniciJessicaNessuna valutazione finora
- Il Sistema Nervoso SemplificatoDocumento5 pagineIl Sistema Nervoso SemplificatoFrancescaNessuna valutazione finora
- Funzioni SensitiveDocumento13 pagineFunzioni SensitiveRoberta RacitiNessuna valutazione finora
- Vie e Circuiti RiverberantiDocumento3 pagineVie e Circuiti RiverberantiOmmalleyNessuna valutazione finora
- Anatomia SNPDocumento30 pagineAnatomia SNPantonio nennaNessuna valutazione finora
- DiencefaloDocumento4 pagineDiencefalocristianNessuna valutazione finora
- Il Sistema NervosoDocumento5 pagineIl Sistema NervosoGianluca LammendolaNessuna valutazione finora
- Funzioni SensitiveDocumento8 pagineFunzioni SensitiveEdoardo BerteroNessuna valutazione finora
- Domande Da Pinel Gennaio 2012Documento13 pagineDomande Da Pinel Gennaio 2012Anna MaranoNessuna valutazione finora
- Domande e Risposte Sistema NervosoDocumento4 pagineDomande e Risposte Sistema Nervosoedup220% (1)
- Sistema NervosoDocumento36 pagineSistema NervosoalinabortaNessuna valutazione finora
- Anatomia Del Sistema NervosoDocumento16 pagineAnatomia Del Sistema NervosoAlysiaNessuna valutazione finora
- Fisiologia Lezione 2Documento5 pagineFisiologia Lezione 2The last chimeraNessuna valutazione finora
- Neuro 2Documento24 pagineNeuro 2Agostino InzerilloNessuna valutazione finora
- Sistema Nervoso Encefalo e Nervi Cranici Capitolo 15Documento27 pagineSistema Nervoso Encefalo e Nervi Cranici Capitolo 15enrico73Nessuna valutazione finora
- La Via Piramidale e La Unità MotoriaDocumento46 pagineLa Via Piramidale e La Unità MotoriaAndrea FerreraNessuna valutazione finora
- Neurofisiologia FilesDocumento13 pagineNeurofisiologia FilesAlessandroE.E.FrancoNessuna valutazione finora
- Anatomia - Lezione 22 (11 - 05 - 2023)Documento4 pagineAnatomia - Lezione 22 (11 - 05 - 2023)Alessandro Maria BaroneNessuna valutazione finora
- Sistema Nervoso Centrale e PerifericoDocumento11 pagineSistema Nervoso Centrale e PerifericoELENA ISABELA TAUTUNessuna valutazione finora
- ENTOMOLOGIADocumento28 pagineENTOMOLOGIAGiorgioNessuna valutazione finora
- 21-04-08 Vie Della Sensibilit+á GeneraleDocumento7 pagine21-04-08 Vie Della Sensibilit+á GeneraleGiovanni VescovoNessuna valutazione finora
- Sistema NervosoDocumento5 pagineSistema NervosoGiuliaNessuna valutazione finora
- Appunti Riabilitazione 2Documento38 pagineAppunti Riabilitazione 2FRANCESCA ROSELLINessuna valutazione finora
- Il Sistema NervosoDocumento3 pagineIl Sistema NervosoMatteo QuintilianiNessuna valutazione finora
- Lezione 19 MacchiarelliDocumento21 pagineLezione 19 Macchiarellilone wolfNessuna valutazione finora
- Tronco Encefalico e BulboDocumento9 pagineTronco Encefalico e BulboGiancarlo AttivissimoNessuna valutazione finora
- Dispensa NeurologiaDocumento14 pagineDispensa NeurologiaDaniele SicilianoNessuna valutazione finora
- Lo Sviluppo della Coordinazione dei Movimenti nell'OntogenesiDa EverandLo Sviluppo della Coordinazione dei Movimenti nell'OntogenesiNessuna valutazione finora
- Algoritmi diagnostico-terapeutici in osteopatiaDa EverandAlgoritmi diagnostico-terapeutici in osteopatiaNessuna valutazione finora
- Endocrinologia - Vet - Anatomia2Documento5 pagineEndocrinologia - Vet - Anatomia2Sabrina CillariNessuna valutazione finora
- Splancnologia - Apparato Digerente-DigestivoDocumento21 pagineSplancnologia - Apparato Digerente-DigestivoSabrina CillariNessuna valutazione finora
- Sistema Nervoso - Occhio e OrecchioDocumento3 pagineSistema Nervoso - Occhio e OrecchioSabrina CillariNessuna valutazione finora
- Anatomia Delle Specie Minori - ConiglioDocumento7 pagineAnatomia Delle Specie Minori - ConiglioSabrina CillariNessuna valutazione finora
- Sistema Nervoso Periferico - Nervi CraniciDocumento10 pagineSistema Nervoso Periferico - Nervi CraniciSabrina CillariNessuna valutazione finora
- Anatomia Delle Specie Minori - UccelliDocumento9 pagineAnatomia Delle Specie Minori - UccelliSabrina CillariNessuna valutazione finora
- Sistema Nervoso CentraleDocumento15 pagineSistema Nervoso CentraleSabrina CillariNessuna valutazione finora
- Osteologia Scheletro AssileDocumento7 pagineOsteologia Scheletro AssileSabrina CillariNessuna valutazione finora
- Mano Del CavalloDocumento5 pagineMano Del CavalloSabrina CillariNessuna valutazione finora
- Osteologia Scheletro AssileDocumento7 pagineOsteologia Scheletro AssileSabrina CillariNessuna valutazione finora
- Osteologia Scheletro AppendicolareDocumento5 pagineOsteologia Scheletro AppendicolareSabrina CillariNessuna valutazione finora
- Mano Del CavalloDocumento5 pagineMano Del CavalloSabrina CillariNessuna valutazione finora
- Miologia VeterinariaDocumento7 pagineMiologia VeterinariaSabrina CillariNessuna valutazione finora
- Art Rolo GiaDocumento4 pagineArt Rolo GiaSabrina CillariNessuna valutazione finora
- Riassunti NeurofisiologiaDocumento25 pagineRiassunti NeurofisiologiaSabrina CillariNessuna valutazione finora
- Osteologia Scheletro AppendicolareDocumento5 pagineOsteologia Scheletro AppendicolareSabrina CillariNessuna valutazione finora
- Osteologia Scheletro AssileDocumento7 pagineOsteologia Scheletro AssileSabrina CillariNessuna valutazione finora
- Apparato Circolatorio SanguignoDocumento7 pagineApparato Circolatorio SanguignoSabrina CillariNessuna valutazione finora
- Osteologia Scheletro AppendicolareDocumento5 pagineOsteologia Scheletro AppendicolareSabrina CillariNessuna valutazione finora
- Apparato Circolatorio LinfaticoDocumento4 pagineApparato Circolatorio LinfaticoSabrina CillariNessuna valutazione finora
- Apparato Circolatorio SanguignoDocumento7 pagineApparato Circolatorio SanguignoSabrina CillariNessuna valutazione finora
- Apparato Circolatorio SanguignoDocumento7 pagineApparato Circolatorio SanguignoSabrina CillariNessuna valutazione finora
- Mano Del CavalloDocumento5 pagineMano Del CavalloSabrina CillariNessuna valutazione finora
- Art Rolo GiaDocumento4 pagineArt Rolo GiaSabrina CillariNessuna valutazione finora
- Art Rolo GiaDocumento4 pagineArt Rolo GiaSabrina CillariNessuna valutazione finora
- Apparato Circolatorio LinfaticoDocumento4 pagineApparato Circolatorio LinfaticoSabrina CillariNessuna valutazione finora
- Anatomia Topografica-Veterinaria-TorinoDocumento2 pagineAnatomia Topografica-Veterinaria-TorinoSabrina CillariNessuna valutazione finora
- Anatomia Topografica-Veterinaria-TorinoDocumento2 pagineAnatomia Topografica-Veterinaria-TorinoSabrina CillariNessuna valutazione finora
- Art Rolo GiaDocumento4 pagineArt Rolo GiaSabrina CillariNessuna valutazione finora
- Preparazione Posizione Del LotoDocumento11 paginePreparazione Posizione Del LotoLuca SiddharthaNessuna valutazione finora
- 4) Sistema VestibolareDocumento9 pagine4) Sistema VestibolareedenNessuna valutazione finora
- Apparato LinfaticoDocumento19 pagineApparato LinfaticoPaolo AnnecchinoNessuna valutazione finora
- Claudia Rainville - METAMEDICINADocumento219 pagineClaudia Rainville - METAMEDICINAHenrique BizarroNessuna valutazione finora
- Neuro 5-Anatomia Descrittiva Del Diencefalo e Del TelencefaloDocumento27 pagineNeuro 5-Anatomia Descrittiva Del Diencefalo e Del TelencefaloRoberto FebeiNessuna valutazione finora
- Anatomia Della PelleDocumento9 pagineAnatomia Della PelleMaria Grazia LopezNessuna valutazione finora
- Clinica OrtopedicaDocumento59 pagineClinica OrtopedicaLuca BevarNessuna valutazione finora
- Muscoli Del RachideDocumento6 pagineMuscoli Del RachideMichele ScipioniNessuna valutazione finora
- Analisi StrutturaleDocumento12 pagineAnalisi StrutturaleDanilo Di VicoNessuna valutazione finora
- L'Apparato EscretoreDocumento2 pagineL'Apparato EscretoreVanny CorteseNessuna valutazione finora
- IRIDOLOGIA Corso BaseDocumento49 pagineIRIDOLOGIA Corso Baseannapolizzotto100% (1)