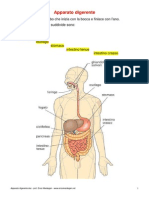
Potrebbero piacerti anche
- Catabolismo AminoacidiDocumento6 pagineCatabolismo AminoacidiDavide BianchiNessuna valutazione finora
- Uomo e NaturaDocumento6 pagineUomo e NaturaAlessia PetragalloNessuna valutazione finora
- Metabolismo Delle ProteineDocumento10 pagineMetabolismo Delle ProteineGianluca Garozzo100% (1)
- Metabolismo Delle ProteineDocumento7 pagineMetabolismo Delle Proteinemauroregazzoni.04Nessuna valutazione finora
- 6 Lezione ScalfiDocumento8 pagine6 Lezione ScalfiClaudio MormoneNessuna valutazione finora
- LEZIONE 12 Metabolismo Dei Composti AzotatiDocumento40 pagineLEZIONE 12 Metabolismo Dei Composti AzotatiMaegazine EnizageamNessuna valutazione finora
- BIOgiustaDocumento29 pagineBIOgiustaguerracristina102Nessuna valutazione finora
- Macronutrienti - Le ProteineDocumento13 pagineMacronutrienti - Le ProteineMario Martin SantoroNessuna valutazione finora
- Glicogeno Lisi e SintesiDocumento3 pagineGlicogeno Lisi e Sintesibenefusco1303Nessuna valutazione finora
- Sintesi Lezioni BiochimicaDocumento156 pagineSintesi Lezioni BiochimicaUmberto Sueri100% (1)
- Amminoacidi e ProteineDocumento64 pagineAmminoacidi e ProteineMajorana.WEBNessuna valutazione finora
- ProteineDocumento2 pagineProteineELEONORA COZZINessuna valutazione finora
- Biochimica EmoglobinaDocumento73 pagineBiochimica EmoglobinaNoemi Di FucciaNessuna valutazione finora
- Il MetabolismoDocumento3 pagineIl MetabolismocuomoidaNessuna valutazione finora
- Biochimica ClinicaDocumento10 pagineBiochimica Clinicablablabla25Nessuna valutazione finora
- Appunti MetabolismoDocumento19 pagineAppunti MetabolismoLuca CanegratiNessuna valutazione finora
- Carboidrati, Proteine e PolisaccaridiDocumento8 pagineCarboidrati, Proteine e PolisaccaridiFederica MellaceNessuna valutazione finora
- Metabolismo Degli AminoacidiDocumento5 pagineMetabolismo Degli AminoacidiPeter OlumataNessuna valutazione finora
- Vitamine IdrosolubileDocumento30 pagineVitamine IdrosolubileMarco GiNessuna valutazione finora
- A1.2d - Farmaci Antiperlipemici (4 - 12-2019)Documento64 pagineA1.2d - Farmaci Antiperlipemici (4 - 12-2019)Massimo PregnolatoNessuna valutazione finora
- Il Metabolismo - AnabolismoDocumento48 pagineIl Metabolismo - AnabolismoCiro LangellaNessuna valutazione finora
- DietologiaDocumento16 pagineDietologiamajholx0% (1)
- Il MetabolismoDocumento5 pagineIl MetabolismoAlice PizziniNessuna valutazione finora
- LEZIONE 9 Ossidazione Acidi GrassiDocumento58 pagineLEZIONE 9 Ossidazione Acidi GrassiMaegazine EnizageamNessuna valutazione finora
- Acidi GrassiDocumento7 pagineAcidi GrassiValentina D'apolloNessuna valutazione finora
- EnzimiDocumento3 pagineEnzimiSimona d'AlessioNessuna valutazione finora
- 11.catabolismo Dei GrassiDocumento59 pagine11.catabolismo Dei GrassiaoooooooooooooNessuna valutazione finora
- Aa e ProteineDocumento18 pagineAa e ProteineAndrea SanzoNessuna valutazione finora
- Apparato DigerenteDocumento32 pagineApparato DigerenteDavide ContiNessuna valutazione finora
- Principi Di Nutrizione e DietologiaDocumento6 paginePrincipi Di Nutrizione e DietologiaAngelo AvellinoNessuna valutazione finora
- BIOCHIMICA - SiliprandiDocumento50 pagineBIOCHIMICA - SiliprandiinfernuxNessuna valutazione finora
- 08 Energia Mitocondri 2020 2021Documento25 pagine08 Energia Mitocondri 2020 2021Alessio BorghigianiNessuna valutazione finora
- Risposte A Domande BiochimicaDocumento7 pagineRisposte A Domande BiochimicaLuigia CentolaNessuna valutazione finora
- Quiz Su AMINOACIDICODocumento7 pagineQuiz Su AMINOACIDICObiagio castronovoNessuna valutazione finora
- Apparato DigerenteDocumento11 pagineApparato DigerenteMary CotugnoNessuna valutazione finora
- 3 Il SangueDocumento9 pagine3 Il Sanguemariannaguadagno00Nessuna valutazione finora
- Tesina Nutrizione Livello Metabolico CarboidratiDocumento15 pagineTesina Nutrizione Livello Metabolico CarboidratiFrancesco IavaroneNessuna valutazione finora
- ProteineDocumento106 pagineProteinetonino883100% (1)
- Lezione 14Documento102 pagineLezione 14yuppisadNessuna valutazione finora
- Il Metabolismo Energetico, Sintesi Del Corso Di Scienze Della TerraDocumento9 pagineIl Metabolismo Energetico, Sintesi Del Corso Di Scienze Della TerraluludevergottiniNessuna valutazione finora
- Appunti MotoriaDocumento5 pagineAppunti Motorialori4669Nessuna valutazione finora
- Proteine Guida ProjectInVictus Rev4Documento37 pagineProteine Guida ProjectInVictus Rev4Paola LuchesNessuna valutazione finora
- Dommande Con Risposta BiochimicaDocumento37 pagineDommande Con Risposta BiochimicayuppisadNessuna valutazione finora
- I LipidiDocumento8 pagineI Lipidimiccgiulia29Nessuna valutazione finora
- 3a Esercitazione LE MACROMOLECOLE PDFDocumento9 pagine3a Esercitazione LE MACROMOLECOLE PDFSimona Di TullioNessuna valutazione finora
- Le ProteineDocumento5 pagineLe ProteineinglesebrexitprogettoNessuna valutazione finora
- HTTPWWW - Scienze.uniroma2.itwp Contentuploads2012034 Proteine Digiuno SNU 2011 PDFDocumento38 pagineHTTPWWW - Scienze.uniroma2.itwp Contentuploads2012034 Proteine Digiuno SNU 2011 PDFNicky TodiscoNessuna valutazione finora
- Biochimica AppuntiDocumento18 pagineBiochimica AppuntiJasmine OliveriNessuna valutazione finora
- Intestino TenueDocumento6 pagineIntestino TenueHghh NgthhNessuna valutazione finora
- 6 LipoproteineDocumento33 pagine6 LipoproteineCate RinaNessuna valutazione finora
- Fondamenti:: CarbonioDocumento13 pagineFondamenti:: CarbonioGreta CaporizziNessuna valutazione finora
- Riassunto BiochimicaDocumento11 pagineRiassunto BiochimicaFrancesco IavaroneNessuna valutazione finora
- I CarboidratiDocumento13 pagineI CarboidratimizarNessuna valutazione finora
- Respirazione e Fermentazione-MicrobiologiaDocumento18 pagineRespirazione e Fermentazione-MicrobiologiaSamuel Di sottoNessuna valutazione finora
- Biochimica Del FegatoDocumento6 pagineBiochimica Del FegatoSalò MarzialiNessuna valutazione finora
- Insulina e GHDocumento8 pagineInsulina e GHNunzia LeoneNessuna valutazione finora
- Biochimica Strutturale PDFDocumento26 pagineBiochimica Strutturale PDFGiulia VizzaNessuna valutazione finora
- Lezione 10 - ValutazioneDocumento5 pagineLezione 10 - ValutazioneAnna De SimoneNessuna valutazione finora
- 2 - La Funzione Energetica Degli AlimentiDocumento15 pagine2 - La Funzione Energetica Degli Alimentipappo pappaNessuna valutazione finora
- EFFETTI COGNITIVI E FISIOLOGICI DELL' ASSUNZIONE DI ACIDI GRASSI POLINSATURI OMEGA-3 IN SOGGETTI SANIDa EverandEFFETTI COGNITIVI E FISIOLOGICI DELL' ASSUNZIONE DI ACIDI GRASSI POLINSATURI OMEGA-3 IN SOGGETTI SANINessuna valutazione finora
- Biochimica II - DomandeDocumento15 pagineBiochimica II - DomandeLorenzo Maria CerviNessuna valutazione finora
- A Metabolismo CellulareDocumento14 pagineA Metabolismo CellularemodenafraNessuna valutazione finora
- Biochimica - Scienze Motorie PDFDocumento282 pagineBiochimica - Scienze Motorie PDFRenatoNessuna valutazione finora
- 05 Fegato 1Documento67 pagine05 Fegato 1Renzo LigurgoNessuna valutazione finora
- Risposte BiochimicaDocumento47 pagineRisposte BiochimicaMarco BiscosiNessuna valutazione finora
- Biochimica Esame 2Documento13 pagineBiochimica Esame 2enzocavajuoloNessuna valutazione finora
- BiochimicDocumento282 pagineBiochimicSalvatore FontanarosaNessuna valutazione finora