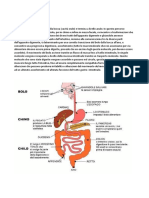
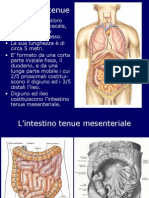
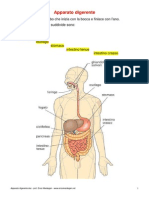
Potrebbero piacerti anche
- Apparato DigerenteDocumento6 pagineApparato DigerenteAnnachiara GargiuloNessuna valutazione finora
- Apparato DigerenteDocumento3 pagineApparato DigerenteGreta SantiNessuna valutazione finora
- Apparato DigerenteDocumento10 pagineApparato DigerenteBruce WiiNessuna valutazione finora
- Digestione E Assorbimento Dei LipidiDocumento30 pagineDigestione E Assorbimento Dei LipidiChiara LassoniNessuna valutazione finora
- Apparato DigerenteDocumento58 pagineApparato DigerenteGiorgio100% (1)
- DietologiaDocumento16 pagineDietologiamajholx0% (1)
- Educazione alimentare. Guida pratica per un'alimentazione sana e naturaleDa EverandEducazione alimentare. Guida pratica per un'alimentazione sana e naturaleNessuna valutazione finora
- DigestioneDocumento11 pagineDigestionesono bruttoNessuna valutazione finora
- Digestione Monogastrici RuminantiDocumento54 pagineDigestione Monogastrici RuminantiEnea VirgilioNessuna valutazione finora
- Sbobina CURS 5Documento5 pagineSbobina CURS 5Giorgia ManieroNessuna valutazione finora
- I LipidiDocumento8 pagineI Lipidimiccgiulia29Nessuna valutazione finora
- Fisiologia Apparato DigerenteDocumento35 pagineFisiologia Apparato Digerentemgrazya89Nessuna valutazione finora
- Elaborato Intestino TenueDocumento7 pagineElaborato Intestino TenueRosaNessuna valutazione finora
- Tesina Nutrizione Livello Metabolico CarboidratiDocumento15 pagineTesina Nutrizione Livello Metabolico CarboidratiFrancesco IavaroneNessuna valutazione finora
- Omeopatia - Lo Stomaco E Le Piante MedicinaliDocumento91 pagineOmeopatia - Lo Stomaco E Le Piante MedicinaliLuca MauraNessuna valutazione finora
- 3.digestione Monogastrici Ruminanti2014Documento60 pagine3.digestione Monogastrici Ruminanti2014Alessandra Patricia LopesNessuna valutazione finora
- L'Apparato DigerenteDocumento13 pagineL'Apparato DigerenteAngelo FernandoNessuna valutazione finora
- Medicina InternaDocumento112 pagineMedicina InternaAutogeneratasiNessuna valutazione finora
- Tratto Gastro IntestinaleDocumento8 pagineTratto Gastro IntestinaleDxneNessuna valutazione finora
- Fisiologia AppuntiDocumento20 pagineFisiologia AppuntienzocavajuoloNessuna valutazione finora
- Lezione 23 Da StamapreDocumento6 pagineLezione 23 Da StamapreMariacarmela MorrealeNessuna valutazione finora
- Biochimica Del FegatoDocumento6 pagineBiochimica Del FegatoSalò MarzialiNessuna valutazione finora
- DigerenteDocumento61 pagineDigerentemarco chiesaNessuna valutazione finora
- L'apparato DigerenteDocumento8 pagineL'apparato DigerentesiriaNessuna valutazione finora
- Apparato DigerenteDocumento104 pagineApparato Digerentesono bruttoNessuna valutazione finora
- 1 Lezione Prof BozzettoDocumento6 pagine1 Lezione Prof BozzettoClaudio MormoneNessuna valutazione finora
- Metabolismo EnergeticoDocumento4 pagineMetabolismo EnergeticoHghh NgthhNessuna valutazione finora
- Apparato DigerenteDocumento7 pagineApparato DigerenteCatalina Valeria MocanuNessuna valutazione finora
- Sistema Digerente: Intestino TenueDocumento24 pagineSistema Digerente: Intestino TenueValentina GambinoNessuna valutazione finora
- Metabolismo Lipidico PDFDocumento44 pagineMetabolismo Lipidico PDFFrancesco SalvadoriNessuna valutazione finora
- Piccolomini Lezione 1Documento13 paginePiccolomini Lezione 1Arian CinigianiNessuna valutazione finora
- Glicogeno Lisi e SintesiDocumento3 pagineGlicogeno Lisi e Sintesibenefusco1303Nessuna valutazione finora
- Apparato Digerente 2Documento15 pagineApparato Digerente 2melissaNessuna valutazione finora
- 15 Apparato Digerente 2Documento14 pagine15 Apparato Digerente 2mariannaguadagno00Nessuna valutazione finora
- Lo Stomaco È Suddiviso in Quattro RegioniDocumento5 pagineLo Stomaco È Suddiviso in Quattro RegioniFranco BonavNessuna valutazione finora
- Apparato DigerenteDocumento5 pagineApparato DigerentegiordacivileNessuna valutazione finora
- Cavazzuti Powerpoint 52075 c6Documento31 pagineCavazzuti Powerpoint 52075 c6ELENA RIVANONessuna valutazione finora
- Lez 3 Fisiologia Umana 2020 2021 Digerente CarducciDocumento47 pagineLez 3 Fisiologia Umana 2020 2021 Digerente CarducciCaterina NuzzoNessuna valutazione finora
- Uomo e NaturaDocumento6 pagineUomo e NaturaAlessia PetragalloNessuna valutazione finora
- 07 Gastroenterologia - 28.10.21Documento24 pagine07 Gastroenterologia - 28.10.21pietropracucciNessuna valutazione finora
- Di Dio-Compiti Scienze NataleDocumento2 pagineDi Dio-Compiti Scienze NataleAlice Di DioNessuna valutazione finora
- LOMBARDO 7 - Il Sistema DigerenteDocumento9 pagineLOMBARDO 7 - Il Sistema Digerentegalluzzo.giuseppemNessuna valutazione finora
- Apparato DigerenteDocumento11 pagineApparato DigerenteMary CotugnoNessuna valutazione finora
- La sindrome dell'intestino pigro per i principianti - Il libro di auto-aiuto - Come interpretare i sintomi dell'intestino pigro, identificare le cause e guarire l'intestinoDa EverandLa sindrome dell'intestino pigro per i principianti - Il libro di auto-aiuto - Come interpretare i sintomi dell'intestino pigro, identificare le cause e guarire l'intestinoNessuna valutazione finora
- Appunti MotoriaDocumento5 pagineAppunti Motorialori4669Nessuna valutazione finora
- Scienze 2Documento7 pagineScienze 2pippoNessuna valutazione finora
- Fisiologia e Fisiopatologia Del Diabete MellitoDocumento10 pagineFisiologia e Fisiopatologia Del Diabete Mellitosiluma11100% (1)
- Biologia CellulareDocumento6 pagineBiologia CellulareClaudia BertenniNessuna valutazione finora
- App. DigerenteDocumento4 pagineApp. DigerenteCarlotta De GrandisNessuna valutazione finora
- Acidi GrassiDocumento7 pagineAcidi GrassiValentina D'apolloNessuna valutazione finora
- Trasporto TubulareDocumento4 pagineTrasporto TubulareHghh NgthhNessuna valutazione finora
- Metabolismo Delle ProteineDocumento10 pagineMetabolismo Delle ProteineGianluca Garozzo100% (1)
- Apparato DigerenteDocumento2 pagineApparato DigerentedavifNessuna valutazione finora
- 11.catabolismo Dei GrassiDocumento59 pagine11.catabolismo Dei GrassiaoooooooooooooNessuna valutazione finora
- BiomolecoleDocumento7 pagineBiomolecolealessioanginiNessuna valutazione finora
- Esocitosi EndocitosiDocumento5 pagineEsocitosi EndocitosiCarlo SchiavoniNessuna valutazione finora
- Apparato DigerenteDocumento9 pagineApparato DigerenteDoara Giulia ChiosaNessuna valutazione finora
- Fisiologia Del Sistema DigestivoDocumento9 pagineFisiologia Del Sistema DigestivoMarcia BalcazarNessuna valutazione finora
- Apparato Digerente pt-2 30 - 11 - 21Documento13 pagineApparato Digerente pt-2 30 - 11 - 21Francesco Spritz MartiniNessuna valutazione finora
- Apparato DigerenteDocumento2 pagineApparato DigerenteFeliciaGuccioneNessuna valutazione finora
- Pancreatite CronicaDocumento23 paginePancreatite CronicaMarco BronziniNessuna valutazione finora
- Emorragie DigestiveDocumento37 pagineEmorragie DigestiveRamiNessuna valutazione finora
- 43-Anatomia II-14.04.2016 - Fegato, Pancreas e MilzaDocumento7 pagine43-Anatomia II-14.04.2016 - Fegato, Pancreas e MilzaSandroNessuna valutazione finora
- Pivot NeurovegetativiDocumento16 paginePivot NeurovegetativiCannata SalvatoreNessuna valutazione finora
- Anatomia Delle Specie Minori - ConiglioDocumento7 pagineAnatomia Delle Specie Minori - ConiglioSabrina CillariNessuna valutazione finora
- Chirurgia 7 Manuale e TeoriaDocumento73 pagineChirurgia 7 Manuale e TeoriaKristi ÇakoNessuna valutazione finora