Potrebbero piacerti anche
- Gas Tri TiDocumento57 pagineGas Tri Tiapi-3733154100% (1)
- Lezione Ormoni GastrointestinaliDocumento28 pagineLezione Ormoni GastrointestinaliAnna De SimoneNessuna valutazione finora
- Digestione Monogastrici RuminantiDocumento54 pagineDigestione Monogastrici RuminantiEnea VirgilioNessuna valutazione finora
- 3.digestione Monogastrici Ruminanti2014Documento60 pagine3.digestione Monogastrici Ruminanti2014Alessandra Patricia LopesNessuna valutazione finora
- Farmaco 1Documento15 pagineFarmaco 1AlessandraManuela VisentinNessuna valutazione finora
- Fisiologia e Fisiopatologia Del Diabete MellitoDocumento10 pagineFisiologia e Fisiopatologia Del Diabete Mellitosiluma11100% (1)
- Sbobina Lezione 15 BiochimicaDocumento15 pagineSbobina Lezione 15 BiochimicaGiorgia de SantisNessuna valutazione finora
- Uomo e NaturaDocumento6 pagineUomo e NaturaAlessia PetragalloNessuna valutazione finora
- Medicina InternaDocumento112 pagineMedicina InternaAutogeneratasiNessuna valutazione finora
- Ormoni Gastrointestinali 2Documento25 pagineOrmoni Gastrointestinali 2Anna De SimoneNessuna valutazione finora
- Amminoacidi e ProteineDocumento64 pagineAmminoacidi e ProteineMajorana.WEBNessuna valutazione finora
- LEZIONE 9 Ossidazione Acidi GrassiDocumento58 pagineLEZIONE 9 Ossidazione Acidi GrassiMaegazine EnizageamNessuna valutazione finora
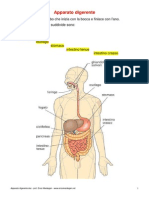
- Apparato DigerenteDocumento10 pagineApparato DigerenteBruce WiiNessuna valutazione finora
- Metabolismo Delle ProteineDocumento10 pagineMetabolismo Delle ProteineGianluca Garozzo100% (1)
- Omeopatia - Lo Stomaco E Le Piante MedicinaliDocumento91 pagineOmeopatia - Lo Stomaco E Le Piante MedicinaliLuca MauraNessuna valutazione finora
- Apparato DigerenteDocumento11 pagineApparato DigerenteMary CotugnoNessuna valutazione finora
- Proteine AlDocumento66 pagineProteine Alrinodelgattopardolibero.itNessuna valutazione finora
- Catabolismo AminoacidiDocumento6 pagineCatabolismo AminoacidiDavide BianchiNessuna valutazione finora
- Med Odonto METABOLISMO DELLE BASI PURINICHE E PIRIMIDINICHEDocumento93 pagineMed Odonto METABOLISMO DELLE BASI PURINICHE E PIRIMIDINICHEFilippo BartoliNessuna valutazione finora
- Coenzimi e VitamineDocumento37 pagineCoenzimi e VitamineRoberta Lo IzzoNessuna valutazione finora
- Riassunto VitamineDocumento7 pagineRiassunto VitamineStefano0% (1)
- Lezione 9Documento54 pagineLezione 9Tonia PreziusoNessuna valutazione finora
- Apparato DigerenteDocumento7 pagineApparato DigerenteCatalina Valeria MocanuNessuna valutazione finora
- 01a Cenni Di Anatomia e Fisiologia Dell'Apparato DigerenteDocumento5 pagine01a Cenni Di Anatomia e Fisiologia Dell'Apparato DigerenteFrancesco NaniaNessuna valutazione finora
- GottaDocumento9 pagineGottaannaricci.aNessuna valutazione finora
- 12 Gastroenterologia PANCREATITIDocumento13 pagine12 Gastroenterologia PANCREATITIangelmartinez87Nessuna valutazione finora
- Metabolismo Calcio FosforoDocumento6 pagineMetabolismo Calcio Fosforothe_answer911572Nessuna valutazione finora
- Combinazioni Alimentari PDFDocumento5 pagineCombinazioni Alimentari PDFmarco poloNessuna valutazione finora
- Biochimica Del FegatoDocumento6 pagineBiochimica Del FegatoSalò MarzialiNessuna valutazione finora
- Patologia Clinica FegatoDocumento14 paginePatologia Clinica FegatoBarbara BranconeNessuna valutazione finora
- A EnzimiDocumento15 pagineA EnzimiBiagio CastronovoNessuna valutazione finora
- DigestioneDocumento11 pagineDigestionesono bruttoNessuna valutazione finora
- Sistema Endocrino e Ormoni Prof. Marco GiammancoDocumento59 pagineSistema Endocrino e Ormoni Prof. Marco GiammancoMarco GiammancoNessuna valutazione finora
- Biochimica AvanzataDocumento15 pagineBiochimica Avanzataapi-3863584100% (3)
- GastroenterologiaDocumento12 pagineGastroenterologianadiaNessuna valutazione finora
- Apparato DigerenteDocumento58 pagineApparato DigerenteGiorgio100% (1)
- LimpidexDocumento99 pagineLimpidexmarioNessuna valutazione finora
- Sbobina CURS 5Documento5 pagineSbobina CURS 5Giorgia ManieroNessuna valutazione finora
- Controllo MetabolismoDocumento31 pagineControllo Metabolismosweet_05Nessuna valutazione finora
- LEZIONE 11 Gluconeogenesi SMODocumento44 pagineLEZIONE 11 Gluconeogenesi SMOMaegazine EnizageamNessuna valutazione finora
- Elenco Quesiti Biochimica ClinicaDocumento10 pagineElenco Quesiti Biochimica ClinicaNicola LauroraNessuna valutazione finora
- Vitamine IdrosolubileDocumento30 pagineVitamine IdrosolubileMarco GiNessuna valutazione finora
- Sistema Gastrointestinale Fisiologia DelDocumento160 pagineSistema Gastrointestinale Fisiologia DelMiriam GalliNessuna valutazione finora
- Apparato DigerenteDocumento104 pagineApparato Digerentesono bruttoNessuna valutazione finora
- 5-Fibra AlimentareDocumento27 pagine5-Fibra Alimentarepappo pappaNessuna valutazione finora
- Biochimica ClinicaDocumento37 pagineBiochimica ClinicaMarco OddeninoNessuna valutazione finora
- 6 Lezione ScalfiDocumento8 pagine6 Lezione ScalfiClaudio MormoneNessuna valutazione finora
- Tesi Roberta Musumeci 9 03 2012Documento78 pagineTesi Roberta Musumeci 9 03 2012api-322196723Nessuna valutazione finora
- Intestino TenueDocumento6 pagineIntestino TenueHghh NgthhNessuna valutazione finora
- 20 Le Aferesi Selettive - TecnologiaDocumento40 pagine20 Le Aferesi Selettive - TecnologiaAndreaNessuna valutazione finora
- Disbiosi Intestinale e Patologie Correlate DR - Lozio LucianoDocumento100 pagineDisbiosi Intestinale e Patologie Correlate DR - Lozio LucianoCosimo Lanza100% (2)
- DietologiaDocumento16 pagineDietologiamajholx0% (1)
- Lezione 23 Da StamapreDocumento6 pagineLezione 23 Da StamapreMariacarmela MorrealeNessuna valutazione finora
- Integrazione Alimetare Nelle Allergie e Intolleranze Alimemtari - Integrazione FunzionaleDocumento66 pagineIntegrazione Alimetare Nelle Allergie e Intolleranze Alimemtari - Integrazione Funzionalemaria maggioNessuna valutazione finora
- 2021 Farmacia Digerente 2 23 26 AprileDocumento78 pagine2021 Farmacia Digerente 2 23 26 AprileChiara ArdauNessuna valutazione finora
- Gastroeneterologia Sunto FinalefDocumento11 pagineGastroeneterologia Sunto FinalefNunzio OlivieroNessuna valutazione finora
- BIOCHIMICA - SiliprandiDocumento50 pagineBIOCHIMICA - SiliprandiinfernuxNessuna valutazione finora
- Tumori SNCDocumento8 pagineTumori SNCNoemi Di FucciaNessuna valutazione finora
- Domande FisiologiaDocumento6 pagineDomande FisiologiaNoemi Di FucciaNessuna valutazione finora
- Lesioni Elementari Della CuteDocumento9 pagineLesioni Elementari Della CuteNoemi Di FucciaNessuna valutazione finora
- Principi Di EmodinamicaDocumento17 paginePrincipi Di EmodinamicaNoemi Di FucciaNessuna valutazione finora
- Semeiotica PancreasDocumento31 pagineSemeiotica PancreasNoemi Di FucciaNessuna valutazione finora
- Elena Vivaldi-Il CantoDocumento25 pagineElena Vivaldi-Il Cantomel martiganNessuna valutazione finora
- Le EmozioniDocumento28 pagineLe EmozioniAnnamaria CosentinoNessuna valutazione finora
- Dispensa DizioneDocumento12 pagineDispensa DizioneStefano CanaliNessuna valutazione finora
- Valori in SangeDocumento4 pagineValori in SangeOzunu DanielNessuna valutazione finora
- Metodo Essentis NazarovDocumento7 pagineMetodo Essentis NazarovPaolaNessuna valutazione finora
- Sistema Immunitario Sprint Pack Pro MedDocumento1 paginaSistema Immunitario Sprint Pack Pro MedAntonio BifulcoNessuna valutazione finora
- Lezione 02 - Neurologia - 2021-2022 Semeiologia Funzione MotoriaDocumento59 pagineLezione 02 - Neurologia - 2021-2022 Semeiologia Funzione Motoriamaria cristina ristagnoNessuna valutazione finora
- Simulazione N°4 Laurea Professioni Sanitarie - Test Di BiologiaDocumento4 pagineSimulazione N°4 Laurea Professioni Sanitarie - Test Di BiologiaMatteo CirelliNessuna valutazione finora