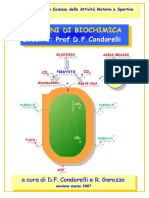
Potrebbero piacerti anche
- Metabolismo Energetico PDFDocumento23 pagineMetabolismo Energetico PDFandrew wallenNessuna valutazione finora
- 122 - Respirazione CellulareDocumento32 pagine122 - Respirazione CellulareFederico Cataldi100% (1)
- Motore Passo PassoDocumento6 pagineMotore Passo PassoGiuseppePeppinoPaciulliNessuna valutazione finora
- Respirazione CellulareDocumento25 pagineRespirazione CellulareLuca GallitelliNessuna valutazione finora
- Dispensa Reality Transurfing Andrea GiavaraDocumento34 pagineDispensa Reality Transurfing Andrea GiavaraAnonymous JBPNLsZJrlNessuna valutazione finora
- Glucose OxidationDocumento38 pagineGlucose OxidationJacks777Nessuna valutazione finora
- Biochimica - Scienze Motorie PDFDocumento282 pagineBiochimica - Scienze Motorie PDFRenatoNessuna valutazione finora
- Metabolismo MicrobicoDocumento19 pagineMetabolismo Microbicodschill2709100% (3)
- Metabolismo - Respirazione Cell. - FotosintesiDocumento10 pagineMetabolismo - Respirazione Cell. - FotosintesiAuroraNessuna valutazione finora
- Metabolismo CellulareDocumento4 pagineMetabolismo CellulareGiada Di GennaroNessuna valutazione finora
- A Metabolismo CellulareDocumento14 pagineA Metabolismo CellularemodenafraNessuna valutazione finora
- Metabolismo CellulareDocumento5 pagineMetabolismo CellulareLeonardo TovoNessuna valutazione finora
- La Respirazione CellulareDocumento2 pagineLa Respirazione CellulareSilvano SpagnoloNessuna valutazione finora
- Ciclo Di KrebsDocumento2 pagineCiclo Di KrebsRosaNessuna valutazione finora
- BioenergeticaDocumento49 pagineBioenergeticaSofia AlvanoNessuna valutazione finora
- Processi Energetici Cellulari 2Documento30 pagineProcessi Energetici Cellulari 2filippo.2003.f8Nessuna valutazione finora
- Il Metabolismo Del GlucosioDocumento3 pagineIl Metabolismo Del GlucosiojolandavigoritaNessuna valutazione finora
- Appunti MetabolismoDocumento19 pagineAppunti MetabolismoLuca CanegratiNessuna valutazione finora
- Cacciatore 09 2017Documento6 pagineCacciatore 09 2017yuppisadNessuna valutazione finora
- Ciclo Di KrebsDocumento5 pagineCiclo Di KrebsCristina StancoNessuna valutazione finora
- Respirazione e Fermentazione-MicrobiologiaDocumento18 pagineRespirazione e Fermentazione-MicrobiologiaSamuel Di sottoNessuna valutazione finora
- La Fosforilazione Ossidativa e La FermentazioneDocumento2 pagineLa Fosforilazione Ossidativa e La FermentazioneCristian ChirNessuna valutazione finora
- Metabolismo EnergeticoDocumento4 pagineMetabolismo EnergeticoMaria Carmela VitielloNessuna valutazione finora
- ChimicaDocumento3 pagineChimicaafaf.mchikhi01Nessuna valutazione finora
- Respirazione CellulareDocumento5 pagineRespirazione CellulareLudovica GaloisNessuna valutazione finora
- Fisiologia VegetaleDocumento8 pagineFisiologia VegetaleChiara IannielloNessuna valutazione finora
- Metabolismo EnergeticoDocumento3 pagineMetabolismo EnergeticoLuca CanegratiNessuna valutazione finora
- Il Metabolismo Dei CarboidratiDocumento7 pagineIl Metabolismo Dei CarboidratiArantxa HoyleNessuna valutazione finora
- Il MetabolismoDocumento5 pagineIl MetabolismoAlice PizziniNessuna valutazione finora
- Ciclo Dell'acido AceticoDocumento5 pagineCiclo Dell'acido AceticoAlessia PapaNessuna valutazione finora
- IL METABOLISMODocumento3 pagineIL METABOLISMOcuomoidaNessuna valutazione finora
- GlicolisiDocumento43 pagineGlicolisigaiamgmalerbaNessuna valutazione finora
- Metabolismo Parte2Documento7 pagineMetabolismo Parte2SimonaNessuna valutazione finora
- Il Fabbisogno Energetico Della CellulaDocumento5 pagineIl Fabbisogno Energetico Della CellulaLuigi BorgeseNessuna valutazione finora
- Il Metabolismo EnergeticoDocumento8 pagineIl Metabolismo EnergeticoPancio BrazzoNessuna valutazione finora
- 24 Respirazione CellulareDocumento1 pagina24 Respirazione CellulareIlaria CribariNessuna valutazione finora
- UntitledDocumento5 pagineUntitledMartina CantelmiNessuna valutazione finora
- Domande BiochimDocumento11 pagineDomande BiochimAlanNessuna valutazione finora
- Risposte A Domande BiochimicaDocumento7 pagineRisposte A Domande BiochimicaLuigia CentolaNessuna valutazione finora
- Il metabolismo energetico, Sintesi del corso di Scienze della terraDocumento9 pagineIl metabolismo energetico, Sintesi del corso di Scienze della terraluludevergottiniNessuna valutazione finora
- GlicolisiDocumento2 pagineGlicolisiCristian UsbergoNessuna valutazione finora
- Respirazione CellulareDocumento29 pagineRespirazione CellulareDaniele ZigiottoNessuna valutazione finora
- La Cellula Al LavoroDocumento36 pagineLa Cellula Al LavoromarioNessuna valutazione finora
- Metabolismo 1Documento46 pagineMetabolismo 1Pippo PlutoNessuna valutazione finora
- FotosintesiDocumento3 pagineFotosintesiLeonardo TovoNessuna valutazione finora
- Ciclo Di KrebsDocumento4 pagineCiclo Di Krebsfox_lgahjzNessuna valutazione finora
- Respirazione_cellulareDocumento28 pagineRespirazione_cellularesiyom81652Nessuna valutazione finora
- ILMETABOLISMOENERGETICODocumento23 pagineILMETABOLISMOENERGETICOdavide abbattistaNessuna valutazione finora
- Sadava Nuova Biologiablu Sintesi A6Documento2 pagineSadava Nuova Biologiablu Sintesi A6Franco 06Nessuna valutazione finora
- 12Documento2 pagine12benedettamari00Nessuna valutazione finora
- Metabolismo Anaerobico Durante Circolazione ExtracorpDocumento46 pagineMetabolismo Anaerobico Durante Circolazione ExtracorpEffetto Di ProssimitàNessuna valutazione finora
- Apparato DigerenteDocumento32 pagineApparato DigerenteDavide ContiNessuna valutazione finora
- Respirazione CellulareDocumento28 pagineRespirazione Cellulareandrew wallenNessuna valutazione finora
- 08 Energia Mitocondri 2020 2021Documento25 pagine08 Energia Mitocondri 2020 2021Alessio BorghigianiNessuna valutazione finora
- Fermentazione LatticaDocumento1 paginaFermentazione LatticaMiriana CostanzoNessuna valutazione finora
- Piante C3 + CLOROFILLADocumento5 paginePiante C3 + CLOROFILLAilsoffionissimoNessuna valutazione finora
- Metabolismo EnergeticoDocumento7 pagineMetabolismo Energeticoxvvdwtbzc9Nessuna valutazione finora
- Uomo e NaturaDocumento6 pagineUomo e NaturaAlessia PetragalloNessuna valutazione finora
- BiochimicaDocumento18 pagineBiochimicajanaNessuna valutazione finora
- Domande di MicrobiologiaDocumento53 pagineDomande di MicrobiologiavalentinodequiNessuna valutazione finora
- Piano Triennale Offerta Formativa 2019 2022Documento2 paginePiano Triennale Offerta Formativa 2019 2022biagio castronovoNessuna valutazione finora
- Verifica Legame Chimico - OcrDocumento3 pagineVerifica Legame Chimico - Ocrbiagio castronovoNessuna valutazione finora
- Invio E-Mail SMARTWATER - White-Paper - Come-avviare-un-impianto-di-depurazione-biologico-Paolo-LunardelliDocumento8 pagineInvio E-Mail SMARTWATER - White-Paper - Come-avviare-un-impianto-di-depurazione-biologico-Paolo-Lunardellibiagio castronovoNessuna valutazione finora
- Piano Intervento AD 20-21Documento14 paginePiano Intervento AD 20-21biagio castronovoNessuna valutazione finora
- Impianti Siracusa Sintesi Non Tecnica VASDocumento289 pagineImpianti Siracusa Sintesi Non Tecnica VASbiagio castronovoNessuna valutazione finora
- Chimica Nei Bienni Degli ITIDocumento6 pagineChimica Nei Bienni Degli ITIbiagio castronovoNessuna valutazione finora
- Libro 1 ADocumento91 pagineLibro 1 Abiagio castronovoNessuna valutazione finora
- CNS 2 2022Documento56 pagineCNS 2 2022biagio castronovoNessuna valutazione finora
- Ciambelle Di PalmaDocumento1 paginaCiambelle Di Palmabiagio castronovoNessuna valutazione finora
- Trattamento Dei Dati AnaliticiDocumento52 pagineTrattamento Dei Dati Analiticibiagio castronovoNessuna valutazione finora
- Distillazione 2Documento24 pagineDistillazione 2Anonymous kTLYjoBqNessuna valutazione finora
- TermochimicaDocumento86 pagineTermochimicabiagio castronovoNessuna valutazione finora
- Esempio Lavoro PoliammidiDocumento17 pagineEsempio Lavoro Poliammidibiagio castronovoNessuna valutazione finora
- Appunti Di Biochimica GeneraleDocumento137 pagineAppunti Di Biochimica Generalebiagio castronovoNessuna valutazione finora
- Manuale Esperienze Laboratorio Chimico BiennioDocumento125 pagineManuale Esperienze Laboratorio Chimico Bienniobiagio castronovo100% (1)
- Legami ChimiciDocumento47 pagineLegami Chimicibiagio castronovoNessuna valutazione finora
- Libro 1 ADocumento91 pagineLibro 1 Abiagio castronovoNessuna valutazione finora
- Appunti Di Chimica Organica PDFDocumento851 pagineAppunti Di Chimica Organica PDFbiagio castronovoNessuna valutazione finora
- Fondamenta Per La Chimica Analitica: Applicazioni Della StatisticaDocumento80 pagineFondamenta Per La Chimica Analitica: Applicazioni Della Statisticabiagio castronovoNessuna valutazione finora
- Tavola Periodica - 1Documento2 pagineTavola Periodica - 1biagio castronovo100% (1)
- Tesi Daniele Zavattini Definitiva NylonDocumento38 pagineTesi Daniele Zavattini Definitiva Nylonbiagio castronovoNessuna valutazione finora
- Biochimica1 IreneCecchiaDocumento81 pagineBiochimica1 IreneCecchiabiagio castronovoNessuna valutazione finora
- Chimica Test Svolti 2017Documento10 pagineChimica Test Svolti 2017biagio castronovoNessuna valutazione finora
- Quiz Su AMINOACIDICODocumento7 pagineQuiz Su AMINOACIDICObiagio castronovoNessuna valutazione finora
- Reattivita 090320 PDFDocumento17 pagineReattivita 090320 PDFbiagio castronovoNessuna valutazione finora
- Appunti Chimica 2 PDFDocumento35 pagineAppunti Chimica 2 PDFbiagio castronovoNessuna valutazione finora
- Amminoacidi e Proteine PDFDocumento33 pagineAmminoacidi e Proteine PDFbiagio castronovoNessuna valutazione finora
- Reattivita 090320Documento17 pagineReattivita 090320biagio castronovoNessuna valutazione finora
- Amminoacidi e Proteine PDFDocumento33 pagineAmminoacidi e Proteine PDFbiagio castronovoNessuna valutazione finora
- 2 AmminoacidiDocumento8 pagine2 Amminoacidibiagio castronovoNessuna valutazione finora
- Raccoltadi Esercizidi ProbabilitaDocumento48 pagineRaccoltadi Esercizidi ProbabilitaJoseph RobertsonNessuna valutazione finora
- FANTONIDocumento13 pagineFANTONIAlejandro Morales0% (1)
- Festival Della Scienza 2013 - Programma PDFDocumento68 pagineFestival Della Scienza 2013 - Programma PDFZadig srlNessuna valutazione finora
- PCB Acetone Al ColDocumento1 paginaPCB Acetone Al ColpippoNessuna valutazione finora