Potrebbero piacerti anche
- Ringiovanire Con Il Plasma Sanguigno Dei GiovaniDa EverandRingiovanire Con Il Plasma Sanguigno Dei GiovaniNessuna valutazione finora
- Citometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheDa EverandCitometria a flusso:principi generali e applicazione allo studio di alcune malattie ematologicheNessuna valutazione finora
- Fisiologia - Fisica Dell'EcgDocumento23 pagineFisiologia - Fisica Dell'EcgBenjaminMeghnagi0% (1)
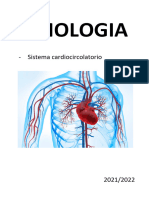
- Fisiologia Del Sistema CardiocircolatorioDocumento175 pagineFisiologia Del Sistema Cardiocircolatorioriky1000Nessuna valutazione finora
- Origine e Propagazione Dell'Eccitamento Nel CuoreDocumento16 pagineOrigine e Propagazione Dell'Eccitamento Nel CuoreRapIoaPer100% (1)
- Fisiologia Degli Apparati (CuoreDocumento146 pagineFisiologia Degli Apparati (CuoreDario Dall'agataNessuna valutazione finora
- MiorisoluzioneDocumento2 pagineMiorisoluzionearabia26Nessuna valutazione finora
- Muscolo LiscioDocumento20 pagineMuscolo Liscioleo.sei.beloNessuna valutazione finora
- FisiologiaDocumento42 pagineFisiologiaLaura TreggiariNessuna valutazione finora
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- Cristal 6Documento32 pagineCristal 6Vale46 RossiNessuna valutazione finora
- 03.ecgmio - Anatomia Vie Di Conduzione e Fisiologia PDFDocumento6 pagine03.ecgmio - Anatomia Vie Di Conduzione e Fisiologia PDFSabrina CenerazzoNessuna valutazione finora
- Biomeccanica Del Tessuto MuscolareDocumento34 pagineBiomeccanica Del Tessuto MuscolareduskyroseNessuna valutazione finora
- FISIOLOGIADocumento6 pagineFISIOLOGIAMariateresa SantorsolaNessuna valutazione finora
- OLIVIERO AUDISIO EstrattoDocumento8 pagineOLIVIERO AUDISIO EstrattoBiancamacarioNessuna valutazione finora
- Cap 4 Riassunto FinDocumento13 pagineCap 4 Riassunto Finandrorru.stefanoNessuna valutazione finora
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- Elettrocardiogramma Tesina Bioingegneria ElettronicaDocumento31 pagineElettrocardiogramma Tesina Bioingegneria ElettronicasmNessuna valutazione finora
- Fisiologia Del ComportamentoDocumento20 pagineFisiologia Del ComportamentoracheleNessuna valutazione finora
- Dispensa Fisiologia UnitoDocumento497 pagineDispensa Fisiologia Unitodorian88Nessuna valutazione finora
- Muscolo ScheletricoDocumento4 pagineMuscolo ScheletricoHghh NgthhNessuna valutazione finora
- Fisiologia Generale, Marco Virgili, Università Di BolognaDocumento165 pagineFisiologia Generale, Marco Virgili, Università Di BolognaSofia PieriNessuna valutazione finora
- (Med ITA) Fisiologia - Automatismo CardiacoDocumento6 pagine(Med ITA) Fisiologia - Automatismo CardiacoZorayda PatriciaNessuna valutazione finora
- Olom-OraDocumento8 pagineOlom-OrariccardosalviNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- Pubblicazioni Sulle MicrocorrentiDocumento13 paginePubblicazioni Sulle Microcorrenticarne23Nessuna valutazione finora
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- Fisiologia PDFDocumento75 pagineFisiologia PDFMarioNessuna valutazione finora
- Fisiologia CardiacaDocumento1 paginaFisiologia CardiacaGaia BorraccesiNessuna valutazione finora
- 2.fisiologia MuscolareDocumento17 pagine2.fisiologia MuscolareHussam SmithNessuna valutazione finora
- AAA Elettroterapia PDFDocumento51 pagineAAA Elettroterapia PDFOsteofisioNessuna valutazione finora
- Muscolo CardiacoDocumento26 pagineMuscolo CardiacomelissaNessuna valutazione finora
- File Completo IstologiaDocumento56 pagineFile Completo IstologiaSara Chiavacci100% (1)
- 14-Emostasi PrimariaDocumento43 pagine14-Emostasi PrimariaSpinelli PietroNessuna valutazione finora
- Sbob Russo-Calabro 2018-2019 PDFDocumento156 pagineSbob Russo-Calabro 2018-2019 PDFStani SantangeloNessuna valutazione finora
- Tessuto MuscolareDocumento3 pagineTessuto Muscolareyeast60Nessuna valutazione finora
- CuoreDocumento48 pagineCuoreLADYOSCARNessuna valutazione finora
- Lezione 3 (13 Ottobre)Documento9 pagineLezione 3 (13 Ottobre)ste MassariNessuna valutazione finora
- Dispositivi Cardiaci ImpiantabiliDocumento101 pagineDispositivi Cardiaci Impiantabilithecorfiot5Nessuna valutazione finora
- Appunti FisiologiaDocumento30 pagineAppunti FisiologiaWartortle BiancanubbaNessuna valutazione finora
- Istologia 42 - Muscolare Liscio PDFDocumento3 pagineIstologia 42 - Muscolare Liscio PDFAli S SaadNessuna valutazione finora
- 22 Lezione Biomeccanica MuscoloDocumento36 pagine22 Lezione Biomeccanica MuscoloFlavio VillaNessuna valutazione finora
- FISIOLOGIADocumento30 pagineFISIOLOGIAlorenzo carliniNessuna valutazione finora
- Fisiologia e Biologia Per Corso Di Ingegneria BiomedicaDocumento13 pagineFisiologia e Biologia Per Corso Di Ingegneria BiomedicaMarco FunaroNessuna valutazione finora
- Il MuscoloDocumento25 pagineIl MuscoloSimone CipollettiNessuna valutazione finora
- Trasporti Di MembranaDocumento10 pagineTrasporti Di MembranaFrancesco BorealeNessuna valutazione finora
- Il Battito CardiacoDocumento4 pagineIl Battito CardiacoGiulia GiuffridaNessuna valutazione finora
- FISIOLOGIA AppuntiDocumento70 pagineFISIOLOGIA AppuntiMarta OreficiniNessuna valutazione finora
- Ematologia Lez 18Documento25 pagineEmatologia Lez 18SIMONA ACANFORANessuna valutazione finora
- Fisiologia UmanaDocumento99 pagineFisiologia UmanaLuis Criscuolo100% (1)
- Attività Elettrica Del CuoreDocumento26 pagineAttività Elettrica Del CuoreStella FilanteNessuna valutazione finora
- Biologia 31 - NucleoDocumento3 pagineBiologia 31 - NucleoFrancesco TarulloNessuna valutazione finora
- Sistema Nervoso EntericoDocumento3 pagineSistema Nervoso EntericophotopemNessuna valutazione finora
- Fisiolgia-Mauro13-Cellule Autoritmiche Ed Innervazione Autonoma Del CuoreDocumento5 pagineFisiolgia-Mauro13-Cellule Autoritmiche Ed Innervazione Autonoma Del CuorecosedasapereNessuna valutazione finora
- 12 Citoscheletro PDFDocumento44 pagine12 Citoscheletro PDFDiego Flores EgüezNessuna valutazione finora
- Verifica Di AnatomiaDocumento3 pagineVerifica Di AnatomiaGelsomino GrilloNessuna valutazione finora
- Tessuto Muscolare PDFDocumento15 pagineTessuto Muscolare PDFLuis CriscuoloNessuna valutazione finora
- MITOCONDRIDocumento4 pagineMITOCONDRIMartina PorcuNessuna valutazione finora
- Conta GlobuliDocumento8 pagineConta Globulimatijacej03speedNessuna valutazione finora
- 1 Potenziale Di Equilibrio Di NernstDocumento5 pagine1 Potenziale Di Equilibrio Di NernstClaudia OrlannoNessuna valutazione finora
- Allergia Sistemica Al Nickel e DietaDocumento24 pagineAllergia Sistemica Al Nickel e DietaSergio Oliva100% (1)
- Vocab Idiomatiche AnimaliDocumento11 pagineVocab Idiomatiche AnimaliPasquale CicchettiNessuna valutazione finora
- Le Malattie Del Legno Della Vite Di Origine Fungina: Salvatorica Serra Renzo PerettoDocumento44 pagineLe Malattie Del Legno Della Vite Di Origine Fungina: Salvatorica Serra Renzo PerettoRoberta PippiNessuna valutazione finora
- Parete Addominale PosterioreDocumento4 pagineParete Addominale Posteriorealessan9Nessuna valutazione finora
- Garoni Il Canile Come Centro Servizi PDFDocumento175 pagineGaroni Il Canile Come Centro Servizi PDFmarinagrossi5212Nessuna valutazione finora