Potrebbero piacerti anche
- Accumulatori per impianti ad energia rinnovabileDa EverandAccumulatori per impianti ad energia rinnovabileValutazione: 2 su 5 stelle2/5 (1)
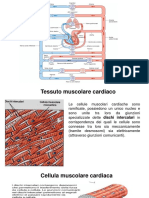
- Attività Elettrica Del CuoreDocumento26 pagineAttività Elettrica Del CuoreStella FilanteNessuna valutazione finora
- Attività Meccanica Del CuoreDocumento7 pagineAttività Meccanica Del CuoreLisaNessuna valutazione finora
- Fisiologia - Contrazione - 2008-2009Documento41 pagineFisiologia - Contrazione - 2008-2009Martina LicchelliNessuna valutazione finora
- Lezione CardiofisiologiaDocumento6 pagineLezione CardiofisiologiaxjaguarkNessuna valutazione finora
- Appunti Di Fisiologia CardiovascolareDocumento17 pagineAppunti Di Fisiologia CardiovascolareTiziana De FeliceNessuna valutazione finora
- Fisiologia CardiacaDocumento1 paginaFisiologia CardiacaGaia BorraccesiNessuna valutazione finora
- 9.segnalazione Cellulare 2 - 2Documento19 pagine9.segnalazione Cellulare 2 - 2Calogero TodaroNessuna valutazione finora
- 14 - Fisiologia - 28-03-2017Documento16 pagine14 - Fisiologia - 28-03-2017SandroNessuna valutazione finora
- Fisiologia II-7 - Il Sistema Di ConduzioneDocumento14 pagineFisiologia II-7 - Il Sistema Di ConduzioneShaun Ivan MuzicNessuna valutazione finora
- Pot CardiaciDocumento47 paginePot CardiaciRiccardo Mason TroiaNessuna valutazione finora
- Proteine Di TrasportoDocumento14 pagineProteine Di Trasportorobertosims007Nessuna valutazione finora
- Teoria SinapticaDocumento5 pagineTeoria SinapticaFran BisoNessuna valutazione finora
- Muscolo ScheletricoDocumento4 pagineMuscolo ScheletricoHghh NgthhNessuna valutazione finora
- Canali Ionici, Fibre NervoseDocumento22 pagineCanali Ionici, Fibre Nervoseapi-3704348100% (1)
- Potenziali D'azioneDocumento5 paginePotenziali D'azioneHghh NgthhNessuna valutazione finora
- Bio Chi MicaDocumento59 pagineBio Chi Micafocaxo5386Nessuna valutazione finora
- Fisiologia Apparato Cardiocircolatorio 1Documento10 pagineFisiologia Apparato Cardiocircolatorio 1Alessandro Riggi0% (1)
- Tessuto MuscolareDocumento3 pagineTessuto Muscolareyeast60Nessuna valutazione finora
- Appunti Fisiologia IDocumento27 pagineAppunti Fisiologia Ikekko88Nessuna valutazione finora
- CanalopatieDocumento37 pagineCanalopatieRiccardo ChecchinNessuna valutazione finora
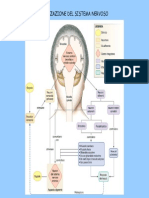
- Il Sistema NervosoDocumento6 pagineIl Sistema NervosoMartina MusmeciNessuna valutazione finora
- Il Sistema NervosoDocumento8 pagineIl Sistema NervosoMartina RossiNessuna valutazione finora
- Fisiologia Del Musculo Cardiaco 2Documento20 pagineFisiologia Del Musculo Cardiaco 2angelfernando2001live.com.mxNessuna valutazione finora
- Scienze 4 VerificaDocumento23 pagineScienze 4 VerificaAlessandra SilveraNessuna valutazione finora
- Sistema NervosoDocumento15 pagineSistema NervosoBruce WiiNessuna valutazione finora
- Muscolo LiscioDocumento14 pagineMuscolo LiscioDaniela PiteoNessuna valutazione finora
- 03.ecgmio - Anatomia Vie Di Conduzione e Fisiologia PDFDocumento6 pagine03.ecgmio - Anatomia Vie Di Conduzione e Fisiologia PDFSabrina CenerazzoNessuna valutazione finora
- Fisiologia Del Sistema CardiocircolatorioDocumento175 pagineFisiologia Del Sistema Cardiocircolatorioriky1000Nessuna valutazione finora
- NEUROBIOLOGIA La SapienzaDocumento30 pagineNEUROBIOLOGIA La Sapienzachiara cimeiNessuna valutazione finora
- Canali Ionici, Enzimi, Molecole CarrierDocumento114 pagineCanali Ionici, Enzimi, Molecole Carrierapi-26155364Nessuna valutazione finora
- Il MuscoloDocumento25 pagineIl MuscoloSimone CipollettiNessuna valutazione finora
- Appunti Modulo 1-2-3Documento84 pagineAppunti Modulo 1-2-3ERIKA BERNOCCHINessuna valutazione finora
- Canali IoniciDocumento3 pagineCanali IoniciFilomena LongobardiNessuna valutazione finora
- Docsity Domande Triennale Fisiologia UmanaDocumento33 pagineDocsity Domande Triennale Fisiologia Umanadestinyema78Nessuna valutazione finora
- Sbob Russo-Calabro 2018-2019 PDFDocumento156 pagineSbob Russo-Calabro 2018-2019 PDFStani SantangeloNessuna valutazione finora
- Pubblicazioni Sulle MicrocorrentiDocumento13 paginePubblicazioni Sulle Microcorrenticarne23Nessuna valutazione finora
- Contrazione Isometrica e IsotonicaDocumento2 pagineContrazione Isometrica e IsotonicaMarco SumaNessuna valutazione finora
- Fisiologia Lezione 4Documento6 pagineFisiologia Lezione 4The last chimeraNessuna valutazione finora
- (PDF - ITA) Manuale Di CardiologiaDocumento352 pagine(PDF - ITA) Manuale Di CardiologiaAlessio Sola75% (8)
- Fisiologia Degli Apparati (CuoreDocumento146 pagineFisiologia Degli Apparati (CuoreDario Dall'agataNessuna valutazione finora
- Accoppiamento Eccitazione-Contrazione Nel CuoreDocumento35 pagineAccoppiamento Eccitazione-Contrazione Nel CuoreLalla BozziNessuna valutazione finora
- Conduzione NervosaDocumento1 paginaConduzione NervosaeurusneNessuna valutazione finora
- Circolazione SanguignaDocumento2 pagineCircolazione Sanguignaao oniNessuna valutazione finora
- Trasmissione Sinaptica e Integrazione NeuronaleDocumento11 pagineTrasmissione Sinaptica e Integrazione NeuronaleAnnamaria MaderaNessuna valutazione finora
- Fisiologia ValenteDocumento4 pagineFisiologia ValenteFrancesco FalconieriNessuna valutazione finora
- 1) Apparato Cardio-VascolareDocumento29 pagine1) Apparato Cardio-Vascolarecor_91Nessuna valutazione finora
- Dispensa Fisiologia UnitoDocumento497 pagineDispensa Fisiologia Unitodorian88Nessuna valutazione finora
- 01-La Genesi Dell'impulso Nervoso PDFDocumento8 pagine01-La Genesi Dell'impulso Nervoso PDFDiego Flores EgüezNessuna valutazione finora
- S NervosoDocumento104 pagineS NervosoAgostino InzerilloNessuna valutazione finora
- Istologia 42 - Muscolare Liscio PDFDocumento3 pagineIstologia 42 - Muscolare Liscio PDFAli S SaadNessuna valutazione finora
- Fisiologia 5 - Fisiologia MuscolareDocumento10 pagineFisiologia 5 - Fisiologia MuscolareAnna De Simone100% (1)
- Fisiologia Generale 5Documento46 pagineFisiologia Generale 5OrlandoCialliNessuna valutazione finora
- Conduzione Nervosa (Riassunto)Documento10 pagineConduzione Nervosa (Riassunto)Aaron RomanoNessuna valutazione finora
- Anche Ad Altre ComponentiDocumento4 pagineAnche Ad Altre Componentifrancesco biancoNessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Sistema CircolatorioDocumento10 pagineSistema CircolatorioLN StylinsonNessuna valutazione finora
- Potenziale D'azioneDocumento30 paginePotenziale D'azioneRiccardo Mason TroiaNessuna valutazione finora
- Sinapsi e NeurotrasmettitoriDocumento45 pagineSinapsi e NeurotrasmettitoriNicola Laurora0% (1)
- Elettrofisiologia CardiacaDocumento8 pagineElettrofisiologia Cardiacaleo.sei.beloNessuna valutazione finora
- MaiPiuReflussoAcido V 17Documento212 pagineMaiPiuReflussoAcido V 17Dada BangNessuna valutazione finora
- GZRic Salsa OlandeseDocumento2 pagineGZRic Salsa Olandesedaki12Nessuna valutazione finora
- DETERMINAZIONICONLOIODI1Documento6 pagineDETERMINAZIONICONLOIODI1rodrigue yanNessuna valutazione finora
- Cei en 60519-1Documento68 pagineCei en 60519-1seve75Nessuna valutazione finora
- Sebastian Faulks - Il Canto Del CieloDocumento235 pagineSebastian Faulks - Il Canto Del CieloAlessandra De AngelisNessuna valutazione finora
- 2 - L'attività SportivaDocumento24 pagine2 - L'attività SportivaMaegazine EnizageamNessuna valutazione finora