Potrebbero piacerti anche
- Introduzione All' IstologiaDocumento6 pagineIntroduzione All' IstologiaLuana LeonardiNessuna valutazione finora
- Pediatria Sbob 20152016Documento321 paginePediatria Sbob 20152016Rosca ValeriuNessuna valutazione finora
- AnatomiaDocumento17 pagineAnatomiaAlice RavasioNessuna valutazione finora
- Soluzioni MariebDocumento7 pagineSoluzioni MariebGabriele DiottiNessuna valutazione finora
- Effetti Somatici Dei Suoni Sistema 1Documento6 pagineEffetti Somatici Dei Suoni Sistema 1Manuel RodriguezNessuna valutazione finora
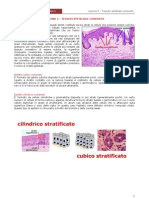
- Lezione 2 - Tessuto Epiteliale CompostoDocumento7 pagineLezione 2 - Tessuto Epiteliale CompostotrillicosaNessuna valutazione finora
- 2 EpiteliRivestimentoDocumento29 pagine2 EpiteliRivestimentoCarolaNessuna valutazione finora
- IstologiaDocumento47 pagineIstologiaSimone Chinaglia0% (1)
- Capitolo 1. L Invecchiamento Della Retina. Introduzione. Organizzazione Morfologica e Funzionale Della Retina PDFDocumento161 pagineCapitolo 1. L Invecchiamento Della Retina. Introduzione. Organizzazione Morfologica e Funzionale Della Retina PDFtopobucciaNessuna valutazione finora
- IstologiaDocumento207 pagineIstologiaVeronica FracassoNessuna valutazione finora
- Introduzione Fasce 2021Documento32 pagineIntroduzione Fasce 2021TomyasuNessuna valutazione finora
- Sbobine Istologia 2021Documento268 pagineSbobine Istologia 2021Au VeNessuna valutazione finora
- TESSUTIDocumento15 pagineTESSUTIRosa NoceraNessuna valutazione finora
- TESSUTIDocumento9 pagineTESSUTIAlice RavasioNessuna valutazione finora
- Differenziamento Cellulare - Derivazione Embrionale IstologiaDocumento17 pagineDifferenziamento Cellulare - Derivazione Embrionale IstologiaWicked_KnightNessuna valutazione finora
- (Dispensa) - ANATOMIA - Ossa, Articolazioni, MuscoliDocumento27 pagine(Dispensa) - ANATOMIA - Ossa, Articolazioni, MuscoliKheper MessoriNessuna valutazione finora
- 2 Anatomia Microscopica e Istologica 2Documento32 pagine2 Anatomia Microscopica e Istologica 2riccardoNessuna valutazione finora
- BiologiaDocumento61 pagineBiologiaElanor OsghiliatNessuna valutazione finora
- Osteopatia Essenziale e tecniche sull'area lombo-pelvicaDa EverandOsteopatia Essenziale e tecniche sull'area lombo-pelvicaNessuna valutazione finora
- Istologia Appunti - OdtDocumento20 pagineIstologia Appunti - Odtprova123100% (1)
- Medicina - Epiteli PDFDocumento110 pagineMedicina - Epiteli PDFArthur GuytonNessuna valutazione finora
- Il Tessuto EpitelialeDocumento6 pagineIl Tessuto EpitelialeDiletta De lucaNessuna valutazione finora
- 3 ConnettivoDocumento57 pagine3 ConnettivoAya El AfiaNessuna valutazione finora
- I TessutiDocumento19 pagineI TessutiGiorgioNessuna valutazione finora
- Wa0063.Documento70 pagineWa0063.merdadiculoNessuna valutazione finora
- IstologiaDocumento72 pagineIstologiajessicagalli.03Nessuna valutazione finora
- Tessuto ConnettivoDocumento3 pagineTessuto ConnettivoCantina Festinger FestingerNessuna valutazione finora
- AnatomiaDocumento81 pagineAnatomias.testellaNessuna valutazione finora
- Istologia - Parte 1 RevisionareDocumento7 pagineIstologia - Parte 1 RevisionareErik AllenatoreNessuna valutazione finora
- Sistema Connettivo Giovanni ChettaDocumento7 pagineSistema Connettivo Giovanni ChettaD. S.Nessuna valutazione finora
- Origine e Propagazione Dell'Eccitamento Nel CuoreDocumento16 pagineOrigine e Propagazione Dell'Eccitamento Nel CuoreRapIoaPer100% (1)
- 02 IstologiaDocumento82 pagine02 IstologiaAlucardNessuna valutazione finora
- Cavazzuti Powerpoint 52075 c2Documento39 pagineCavazzuti Powerpoint 52075 c2pincoNessuna valutazione finora
- Testo - Riequilibrio Fasciale, Approccio OsteopaticoDocumento9 pagineTesto - Riequilibrio Fasciale, Approccio Osteopaticoconnective.promoNessuna valutazione finora
- CitologiaDocumento14 pagineCitologiaTitti TeleseNessuna valutazione finora
- 4 5 Tegumento e LocomotoreDocumento20 pagine4 5 Tegumento e LocomotoreELENA ISABELA TAUTUNessuna valutazione finora
- Cellula ProcarioteDocumento3 pagineCellula ProcarioteCarlo Edoardo CapponiNessuna valutazione finora
- Struttura Primaria Del FustoDocumento6 pagineStruttura Primaria Del FustoAndrea MaselliNessuna valutazione finora
- Le Cellule Eucariotiche - BookinprogressDocumento12 pagineLe Cellule Eucariotiche - BookinprogressCarmenNessuna valutazione finora
- Stecco 00Documento14 pagineStecco 00Dandino SpellmanNessuna valutazione finora
- 9-10 Tessusto Epiteliale + Sistema EndocrinoDocumento12 pagine9-10 Tessusto Epiteliale + Sistema Endocrinomariannaguadagno00Nessuna valutazione finora
- MuscolareDocumento15 pagineMuscolareEugenio StracuzzaNessuna valutazione finora
- Istologia Ps Classi 2,3,4 2019-2020Documento304 pagineIstologia Ps Classi 2,3,4 2019-2020Matteo RussoNessuna valutazione finora
- Cellula VegetaDocumento125 pagineCellula VegetaRIGOR dooNessuna valutazione finora
- I Tessuti VegetaliDocumento13 pagineI Tessuti Vegetalialice9980Nessuna valutazione finora
- Appunti CusellaDocumento42 pagineAppunti Cusellalorenzo carliniNessuna valutazione finora
- Lezione 3 - Tessuto Epiteliale Sensoriale e Ghiandolare EsocrinoDocumento5 pagineLezione 3 - Tessuto Epiteliale Sensoriale e Ghiandolare EsocrinotrillicosaNessuna valutazione finora
- Power Point Tessuti BiologiaDocumento11 paginePower Point Tessuti BiologiaRebecca VerdeNessuna valutazione finora
- Biologia 27 - Membrana Cellulare - AdesioneDocumento3 pagineBiologia 27 - Membrana Cellulare - AdesioneFrancesco TarulloNessuna valutazione finora
- Citoscheletro, Matrice Extracellulare e Giunzioni CellulariDocumento37 pagineCitoscheletro, Matrice Extracellulare e Giunzioni CellulariVidabegNessuna valutazione finora
- La Matrice EXTRACELLULAREDocumento5 pagineLa Matrice EXTRACELLULAREManuel RodriguezNessuna valutazione finora
- MiofascialeDocumento7 pagineMiofascialePippoNessuna valutazione finora
- Dermatologia, Venerologia, Chirurgia PlasticaDocumento124 pagineDermatologia, Venerologia, Chirurgia PlasticaMorena Saudino CeisNessuna valutazione finora
- Lezione 1 - Tessuto Epiteliale SempliceDocumento4 pagineLezione 1 - Tessuto Epiteliale Semplicetrillicosa100% (1)
- Lezione 1-2Documento29 pagineLezione 1-2salvatore palmieriNessuna valutazione finora
- FoschiniDocumento42 pagineFoschiniDario Dall'agataNessuna valutazione finora
- Biochimica AppuntiDocumento146 pagineBiochimica AppuntiAlessia Mandorino0% (1)
- Parete OrganuliDocumento54 pagineParete OrganuliAngelo TranoNessuna valutazione finora
- BILOGIA - Tessuti AnimaliDocumento7 pagineBILOGIA - Tessuti AnimaliAndrea ColomboNessuna valutazione finora
- Fisiologia VestibolareDocumento14 pagineFisiologia VestibolareAlinaNessuna valutazione finora
- DOMANDEDocumento9 pagineDOMANDEClaire ColomboNessuna valutazione finora
- LgdisfagiaDocumento41 pagineLgdisfagiaDomenico TurcoNessuna valutazione finora
- Apparato Di Risonanza e Di ArticolazioneDocumento12 pagineApparato Di Risonanza e Di ArticolazioneClaire ColomboNessuna valutazione finora
- AFASIADocumento120 pagineAFASIAClaire ColomboNessuna valutazione finora
- Apparato Di Risonanza e Di ArticolazioneDocumento12 pagineApparato Di Risonanza e Di ArticolazioneClaire ColomboNessuna valutazione finora
- Esame Spettroacustico CasolinoDocumento37 pagineEsame Spettroacustico CasolinoClaire ColomboNessuna valutazione finora
- EmbriologiaDocumento7 pagineEmbriologiaClaire ColomboNessuna valutazione finora
- IstologiaDocumento35 pagineIstologiaClaire ColomboNessuna valutazione finora
- Domande MadedduDocumento13 pagineDomande MadedduClaire ColomboNessuna valutazione finora
- Lezione 2Documento26 pagineLezione 2Claire ColomboNessuna valutazione finora
- Domande DemoetnoantropologiaDocumento1 paginaDomande DemoetnoantropologiaClaire ColomboNessuna valutazione finora
- Lezione 1Documento27 pagineLezione 1Claire ColomboNessuna valutazione finora
- Domande Di LinguisticaDocumento2 pagineDomande Di LinguisticaClaire ColomboNessuna valutazione finora
- MatematicaDocumento10 pagineMatematicaClaire ColomboNessuna valutazione finora
- Planning TestDocumento2 paginePlanning TestClaire ColomboNessuna valutazione finora
- DIURETICIDocumento66 pagineDIURETICIακηςNessuna valutazione finora
- G Rondelli Il Fartlek PIDocumento6 pagineG Rondelli Il Fartlek PIDemetrio FrattarelliNessuna valutazione finora
- Domande e Risposte ARGIOLASDocumento23 pagineDomande e Risposte ARGIOLASDrumsoNessuna valutazione finora
- Il Massaggio e Le Sue Tecniche PDFDocumento24 pagineIl Massaggio e Le Sue Tecniche PDFladydaladyNessuna valutazione finora
- Apparato Muscolare DietroDocumento1 paginaApparato Muscolare DietroLallo MoraniNessuna valutazione finora
- Terapie Fisiche in Ortopedia (Dott. Corrado Greco)Documento54 pagineTerapie Fisiche in Ortopedia (Dott. Corrado Greco)Giorgio Parisi3Nessuna valutazione finora