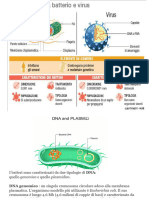
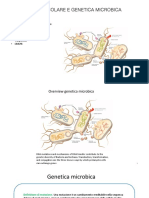
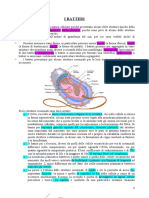
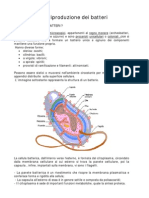
Potrebbero piacerti anche
- Verso La Costruzione Di Vettori Per Il Trasferimento Genico Negli InvertebratiDa EverandVerso La Costruzione Di Vettori Per Il Trasferimento Genico Negli InvertebratiNessuna valutazione finora
- Capitolo 4 GeneticaDocumento4 pagineCapitolo 4 GeneticaMartina CantelmiNessuna valutazione finora
- L’alcaloide vegetale Voacamina esercita un effetto chemiosensibilizzante su cellule tumorali farmacoresistenti mediante l’induzione del processo autofagicoDa EverandL’alcaloide vegetale Voacamina esercita un effetto chemiosensibilizzante su cellule tumorali farmacoresistenti mediante l’induzione del processo autofagicoNessuna valutazione finora
- La Genetiica Di Virus e BatteriDocumento5 pagineLa Genetiica Di Virus e BatteriTimoteo Raffaele RizzatoNessuna valutazione finora
- La Ricombinazione Genica Nei BatteriDocumento5 pagineLa Ricombinazione Genica Nei BatteriVincenzo Di GioiaNessuna valutazione finora
- Virus Regolazioni Procarioti Ed EucariotiDocumento3 pagineVirus Regolazioni Procarioti Ed EucariotiGianmarco VindigniNessuna valutazione finora
- Riassunto EpigeneticaDocumento6 pagineRiassunto EpigeneticaMari 014Nessuna valutazione finora
- 6 MEDICINA - Genetica BattericaDocumento46 pagine6 MEDICINA - Genetica BattericaClaudio GarziaNessuna valutazione finora
- Genetica BacterianaDocumento4 pagineGenetica BacterianaSantiago Franco Eunice GetzemaníNessuna valutazione finora
- Ing GeneticaDocumento49 pagineIng Geneticafernando MartinNessuna valutazione finora
- Virus e BatteriDocumento1 paginaVirus e BatterifarloccoloccoNessuna valutazione finora
- FagiDocumento4 pagineFagiAlexNessuna valutazione finora
- MICROBIOLOGIA Prof Scalia Università Di CataniaDocumento19 pagineMICROBIOLOGIA Prof Scalia Università Di CataniaLucia CasellaNessuna valutazione finora
- Riassunti BonattiDocumento36 pagineRiassunti Bonattiapi-3863584100% (1)
- Virus e Batteri ZanichelliDocumento17 pagineVirus e Batteri Zanichelliandrea.daloisio2007Nessuna valutazione finora
- Organica BiochimicaDocumento12 pagineOrganica BiochimicaDaniel GrabNessuna valutazione finora
- MicrobiologiaDocumento9 pagineMicrobiologiaMarinela GiciuNessuna valutazione finora
- Genetica Microbica 1 Trasformazione-Trasduzione PDFDocumento33 pagineGenetica Microbica 1 Trasformazione-Trasduzione PDFFederica ZanellaNessuna valutazione finora
- 45 Patologia TumorSoppressorGenes Volinia 10 12 2020Documento16 pagine45 Patologia TumorSoppressorGenes Volinia 10 12 2020raffaele.bianchi1997Nessuna valutazione finora
- BatteriDocumento11 pagineBatteriantonella vitaleNessuna valutazione finora
- Riassunti Terapia Genica - Doc 1Documento29 pagineRiassunti Terapia Genica - Doc 1cirocandidoNessuna valutazione finora
- Miv Sistemi Di SecrezioneDocumento5 pagineMiv Sistemi Di Secrezioneantonio nennaNessuna valutazione finora
- Genetica in Batteri e BatteriofagiDocumento56 pagineGenetica in Batteri e BatteriofagiEleonora AngelocolaNessuna valutazione finora
- MICROORGANISMIDocumento3 pagineMICROORGANISMILauraNessuna valutazione finora
- Microbiologia Lezione 4Documento12 pagineMicrobiologia Lezione 4Erica ColellaNessuna valutazione finora
- Elementi TrasponibiliDocumento2 pagineElementi TrasponibiliMiriam CartaNessuna valutazione finora
- Microbiologia 21 Aprile 2020Documento18 pagineMicrobiologia 21 Aprile 2020lorellaNessuna valutazione finora
- VirologiaDocumento64 pagineVirologiaFilippoNessuna valutazione finora
- Batteri, AppuntiDocumento12 pagineBatteri, AppuntiMatteo PeruzzoNessuna valutazione finora
- Appunti Di Tecnica Produzione AnimaleDocumento152 pagineAppunti Di Tecnica Produzione AnimaleDanyfive100% (1)
- Micro InsiemeDocumento39 pagineMicro InsiemeCaterina SarnicolaNessuna valutazione finora
- Procarioti e VirusDocumento14 pagineProcarioti e Virusgiorgio.manfredi.04Nessuna valutazione finora
- BATTERI2Documento5 pagineBATTERI2davideNessuna valutazione finora
- Bio Medico-Diagnostiche 4Documento9 pagineBio Medico-Diagnostiche 4Simona LeopardiNessuna valutazione finora
- Terapia GenicaDocumento31 pagineTerapia GenicagiovanniNessuna valutazione finora
- L5 StrutturaDellaCellulaDocumento42 pagineL5 StrutturaDellaCellulaterezaNessuna valutazione finora
- I BatteriofagiDocumento4 pagineI BatteriofagiLuigi PascaleNessuna valutazione finora
- Lez 1 MicroDocumento6 pagineLez 1 Microclacelove2004Nessuna valutazione finora
- Ciclo Cellulare e Replicazione Del DNADocumento6 pagineCiclo Cellulare e Replicazione Del DNAGiovanni SciaccoNessuna valutazione finora
- BiotecnologieDocumento5 pagineBiotecnologieDelia GuidaNessuna valutazione finora
- Filogenesi Molecolare 1Documento7 pagineFilogenesi Molecolare 1STEFANO BORDIGNONNessuna valutazione finora
- Virus MicrobiologiaDocumento25 pagineVirus MicrobiologiaSamuel Di sottoNessuna valutazione finora
- Lezione 02-FisiologiaDocumento5 pagineLezione 02-FisiologiaSarah ScuderiNessuna valutazione finora
- Riassunto ClonaggioDocumento7 pagineRiassunto ClonaggioDaniela VerrastroNessuna valutazione finora
- 6 Meiosi e Cicli SessualiDocumento17 pagine6 Meiosi e Cicli SessualiAlessia MaraNessuna valutazione finora
- Comunicazione CellulareDocumento8 pagineComunicazione CellularePulciNellerNessuna valutazione finora
- Batter IDocumento38 pagineBatter IMatilde VeronelliNessuna valutazione finora
- Appunti Biologia Molecolare e Evoluzione Dei GenomiDocumento9 pagineAppunti Biologia Molecolare e Evoluzione Dei Genomidsofia2001Nessuna valutazione finora
- Botanica BatteriDocumento4 pagineBotanica BatteriAdriana PellitteriNessuna valutazione finora
- Organismi e MicrorganismiDocumento3 pagineOrganismi e MicrorganismiSimoneNessuna valutazione finora
- Bio Tec No LogieDocumento3 pagineBio Tec No LogieantoniettaNessuna valutazione finora
- 6) Streptococchi e StafilococchiDocumento7 pagine6) Streptococchi e StafilococchibathildaNessuna valutazione finora
- La Neurofibromatosi - Tipo 2Documento51 pagineLa Neurofibromatosi - Tipo 2caila8716Nessuna valutazione finora
- HerpesvirusDocumento103 pagineHerpesvirusFrancesco ScorzaNessuna valutazione finora
- Domande Esame MicrobiologiaDocumento5 pagineDomande Esame MicrobiologiamichNessuna valutazione finora
- Docsity Meccanismi EpigeneticiDocumento7 pagineDocsity Meccanismi EpigeneticiCaterina ZaninNessuna valutazione finora
- Trascrizione e Traduzione Nei ProcariotiDocumento2 pagineTrascrizione e Traduzione Nei ProcariotiClaudia BertenniNessuna valutazione finora
- A1 Origine Ed Evoluzione Delle CelluleDocumento3 pagineA1 Origine Ed Evoluzione Delle CelluleAnna ForasiepiNessuna valutazione finora
- Appunti Di Farmacologia SpecialeDocumento96 pagineAppunti Di Farmacologia SpecialeStefano Vezza100% (1)
- 5 Ciclo Cellulare e MitosiDocumento22 pagine5 Ciclo Cellulare e MitosiAlessia MaraNessuna valutazione finora
- Crop Circles - Interpretazione Achetipica (Corrado Malanga)Documento36 pagineCrop Circles - Interpretazione Achetipica (Corrado Malanga)Angel Kayn100% (3)
- Azian Nicola 2011Documento106 pagineAzian Nicola 2011MarcoCarcano100% (1)
- Campo MorficoDocumento9 pagineCampo Morficoevia311Nessuna valutazione finora
- Riassunto GeneticaDocumento20 pagineRiassunto Geneticamonta89Nessuna valutazione finora
- Istologia CompletaDocumento333 pagineIstologia CompletaCinzia Ciervo67% (3)
- Riassunti Terapia Genica - Doc 1Documento29 pagineRiassunti Terapia Genica - Doc 1cirocandidoNessuna valutazione finora
- Quiz Ammissione Biologia Prima ParteDocumento69 pagineQuiz Ammissione Biologia Prima Parterodrigue yanNessuna valutazione finora
- Capitoli B3 e B4Documento11 pagineCapitoli B3 e B4Gabriele PalermoNessuna valutazione finora
- Trucchi psicologici: Tecniche di comunicazione e programmazione neurolinguistica applicate al mentalismo modernoDa EverandTrucchi psicologici: Tecniche di comunicazione e programmazione neurolinguistica applicate al mentalismo modernoValutazione: 3 su 5 stelle3/5 (2)
- Nomofobia: Cos'è, chi ne soffre, perché esisteDa EverandNomofobia: Cos'è, chi ne soffre, perché esisteNessuna valutazione finora
- La musicoterapia quale didattica applicata ai disturbi dello spettro autisticoDa EverandLa musicoterapia quale didattica applicata ai disturbi dello spettro autisticoNessuna valutazione finora
- Prevenire le Malattie: Strategie Rivoluzionarie per un Domani più Sano: Potenziare il Tuo Percorso verso il Benessere attraverso la Scienza e l'InnovazioneDa EverandPrevenire le Malattie: Strategie Rivoluzionarie per un Domani più Sano: Potenziare il Tuo Percorso verso il Benessere attraverso la Scienza e l'InnovazioneNessuna valutazione finora
- Depressione, ansia e panico: la terapia psicologicaDa EverandDepressione, ansia e panico: la terapia psicologicaNessuna valutazione finora
- The Secret of Spirit-Thought Magic - Now-Project the thoughts that summon spirits from the Magic Astral World to make your secret wishes come true!Da EverandThe Secret of Spirit-Thought Magic - Now-Project the thoughts that summon spirits from the Magic Astral World to make your secret wishes come true!Valutazione: 5 su 5 stelle5/5 (6)
- La teoria della terra cava. The hollow earth theoryDa EverandLa teoria della terra cava. The hollow earth theoryValutazione: 3 su 5 stelle3/5 (1)
- Biologia quantica: Viaggio ai confini della guarigioneDa EverandBiologia quantica: Viaggio ai confini della guarigioneNessuna valutazione finora
- Siamo tutti Cardiopatici: Aumentare l'efficienza cardiovascolareDa EverandSiamo tutti Cardiopatici: Aumentare l'efficienza cardiovascolareNessuna valutazione finora
- Come realizzare cose meravigliose … in un mondo di merda!: Metodi pratici per crescere, realizzare, cambiare le cose... ... anche quando tutto il mondo ci rema contro!Da EverandCome realizzare cose meravigliose … in un mondo di merda!: Metodi pratici per crescere, realizzare, cambiare le cose... ... anche quando tutto il mondo ci rema contro!Nessuna valutazione finora
- Demenza senile e rigenerazione del cervello: Rimanere mentalmente giovani nonostante l’etàDa EverandDemenza senile e rigenerazione del cervello: Rimanere mentalmente giovani nonostante l’etàNessuna valutazione finora
- Esercizi svolti di Matematica: Geometria AnaliticaDa EverandEsercizi svolti di Matematica: Geometria AnaliticaNessuna valutazione finora
- Agnihotri - I Custodi del Fuoco: Il Sentiero di Guarigione del Guerriero del FuocoDa EverandAgnihotri - I Custodi del Fuoco: Il Sentiero di Guarigione del Guerriero del FuocoNessuna valutazione finora
- Linfedema: Spunti pratici per la terapia multidisciplinareDa EverandLinfedema: Spunti pratici per la terapia multidisciplinareNessuna valutazione finora
- Sconfiggere la stanchezza cronica: come riprendersi la propria vitaDa EverandSconfiggere la stanchezza cronica: come riprendersi la propria vitaNessuna valutazione finora
- Ricettario della Paleodieta Autoimmune Le Migliori 30 Ricette della Paleodieta Autoimmune!Da EverandRicettario della Paleodieta Autoimmune Le Migliori 30 Ricette della Paleodieta Autoimmune!Nessuna valutazione finora
- Il trattamento fasciale osteopatico per il sistema nervoso centrale e perifericoDa EverandIl trattamento fasciale osteopatico per il sistema nervoso centrale e perifericoValutazione: 5 su 5 stelle5/5 (1)