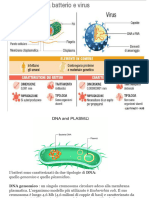
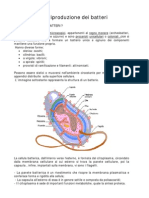
Potrebbero piacerti anche
- Jules Verne - La Scuola Dei RobinsonDocumento188 pagineJules Verne - La Scuola Dei Robinsonfulvix88Nessuna valutazione finora
- sintesiCap4Saraceni 2012 PDFDocumento9 paginesintesiCap4Saraceni 2012 PDFterezaNessuna valutazione finora
- Virus e BatteriDocumento1 paginaVirus e BatterifarloccoloccoNessuna valutazione finora
- Riassunto EpigeneticaDocumento6 pagineRiassunto EpigeneticaMari 014Nessuna valutazione finora
- La Genetiica Di Virus e BatteriDocumento5 pagineLa Genetiica Di Virus e BatteriTimoteo Raffaele RizzatoNessuna valutazione finora
- Virus (Struttura e Replicazione)Documento4 pagineVirus (Struttura e Replicazione)Maria SarbashNessuna valutazione finora
- I Diversi Tipi Di Virus RICERCA SCUOLA SCIENZEDocumento6 pagineI Diversi Tipi Di Virus RICERCA SCUOLA SCIENZEglorieta asmariNessuna valutazione finora
- Organica BiochimicaDocumento12 pagineOrganica BiochimicaDaniel GrabNessuna valutazione finora
- VirologiaDocumento64 pagineVirologiaFilippoNessuna valutazione finora
- Capitolo 4 GeneticaDocumento4 pagineCapitolo 4 GeneticaMartina CantelmiNessuna valutazione finora
- I VirusDocumento6 pagineI VirusValerio MastrapasquaNessuna valutazione finora
- I VirusDocumento5 pagineI VirusSofia CannovaNessuna valutazione finora
- VirologiaDocumento16 pagineVirologiasweet_05Nessuna valutazione finora
- Virus MicrobiologiaDocumento25 pagineVirus MicrobiologiaSamuel Di sottoNessuna valutazione finora
- VIROLOGIADocumento39 pagineVIROLOGIAnacarannaNessuna valutazione finora
- Citologia ?-ConvertitoDocumento77 pagineCitologia ?-ConvertitoEdithNessuna valutazione finora
- Microbiologia Lezione 4Documento12 pagineMicrobiologia Lezione 4Erica ColellaNessuna valutazione finora
- La Ricombinazione Genica Nei BatteriDocumento5 pagineLa Ricombinazione Genica Nei BatteriVincenzo Di GioiaNessuna valutazione finora
- Trascrizione e Traduzione Nei ProcariotiDocumento2 pagineTrascrizione e Traduzione Nei ProcariotiClaudia BertenniNessuna valutazione finora
- Virus INDocumento26 pagineVirus INLapo MosettiNessuna valutazione finora
- MicrobiologiaDocumento9 pagineMicrobiologiaMarinela GiciuNessuna valutazione finora
- Biologia Applicata - VirusDocumento27 pagineBiologia Applicata - Virusapi-3826003100% (3)
- Riassunto ClonaggioDocumento7 pagineRiassunto ClonaggioDaniela VerrastroNessuna valutazione finora
- FagiDocumento4 pagineFagiAlexNessuna valutazione finora
- Genetica in Batteri e BatteriofagiDocumento56 pagineGenetica in Batteri e BatteriofagiEleonora AngelocolaNessuna valutazione finora
- Virus e PandemieDocumento8 pagineVirus e PandemieGianmarco VindigniNessuna valutazione finora
- Terapia GenicaDocumento31 pagineTerapia GenicagiovanniNessuna valutazione finora
- Lez 10 - Virus PDFDocumento52 pagineLez 10 - Virus PDFabyutzaNessuna valutazione finora
- Verso La Costruzione Di Vettori Per Il Trasferimento Genico Negli InvertebratiDa EverandVerso La Costruzione Di Vettori Per Il Trasferimento Genico Negli InvertebratiNessuna valutazione finora
- VirusDocumento3 pagineVirusTHE KILLERNessuna valutazione finora
- Procarioti e VirusDocumento14 pagineProcarioti e Virusgiorgio.manfredi.04Nessuna valutazione finora
- Clonaggio GenicoDocumento4 pagineClonaggio GenicoGreta Video utiliNessuna valutazione finora
- VirusDocumento7 pagineVirusAnna MancaNessuna valutazione finora
- HerpesvirusDocumento103 pagineHerpesvirusFrancesco ScorzaNessuna valutazione finora
- Tecniche MolecolariDocumento8 pagineTecniche Molecolaripewama5294Nessuna valutazione finora
- I VirusDocumento2 pagineI VirusFabio LibertàNessuna valutazione finora
- VirologiaDocumento14 pagineVirologiaSISNessuna valutazione finora
- VirologiaDocumento66 pagineVirologiaAntonio BleveNessuna valutazione finora
- Tgo MicrobiologiaDocumento32 pagineTgo MicrobiologiaSamuel Di sottoNessuna valutazione finora
- Virus e Batteri ZanichelliDocumento17 pagineVirus e Batteri Zanichelliandrea.daloisio2007Nessuna valutazione finora
- 21 MaggioDocumento8 pagine21 MaggiostefanoNessuna valutazione finora
- Genetica BacterianaDocumento4 pagineGenetica BacterianaSantiago Franco Eunice GetzemaníNessuna valutazione finora
- VirologiaDocumento151 pagineVirologiaSimoiadeNessuna valutazione finora
- BiotecnologieDocumento5 pagineBiotecnologieDelia GuidaNessuna valutazione finora
- BATTERI2Documento5 pagineBATTERI2davideNessuna valutazione finora
- Ing GeneticaDocumento49 pagineIng Geneticafernando MartinNessuna valutazione finora
- ParassitologiaDocumento26 pagineParassitologiaSpinelli PietroNessuna valutazione finora
- Elementi TrasponibiliDocumento2 pagineElementi TrasponibiliMiriam CartaNessuna valutazione finora
- Bio Medico-Diagnostiche 4Documento9 pagineBio Medico-Diagnostiche 4Simona LeopardiNessuna valutazione finora
- I VirusDocumento5 pagineI VirusLaura CateniNessuna valutazione finora
- Ciclo CellulareDocumento28 pagineCiclo CellulareMarco BronziniNessuna valutazione finora
- Botanica BatteriDocumento4 pagineBotanica BatteriAdriana PellitteriNessuna valutazione finora
- BIOTECNOLOGIEDocumento5 pagineBIOTECNOLOGIEAlessandro IsidoriNessuna valutazione finora
- Test Biooo Aa ZZZDocumento11 pagineTest Biooo Aa ZZZIlisa SpahiuNessuna valutazione finora
- MICROBIOLOGIA Prof Scalia Università Di CataniaDocumento19 pagineMICROBIOLOGIA Prof Scalia Università Di CataniaLucia CasellaNessuna valutazione finora
- I BatteriofagiDocumento4 pagineI BatteriofagiLuigi PascaleNessuna valutazione finora
- Lezione 2-11-09Documento77 pagineLezione 2-11-09simo_pNessuna valutazione finora
- Lez 3 MicroDocumento12 pagineLez 3 Microclacelove2004Nessuna valutazione finora
- Domande Esame MicrobiologiaDocumento5 pagineDomande Esame MicrobiologiamichNessuna valutazione finora
- Ciclo Cellulare e Replicazione Del DNADocumento6 pagineCiclo Cellulare e Replicazione Del DNAGiovanni SciaccoNessuna valutazione finora
- Morte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cDa EverandMorte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cNessuna valutazione finora
- Genetica Mendeliana e Duplicazione DNADocumento10 pagineGenetica Mendeliana e Duplicazione DNAGianmarco VindigniNessuna valutazione finora
- Schemi MendelDocumento8 pagineSchemi MendelGianmarco VindigniNessuna valutazione finora
- Divisione CellulareDocumento6 pagineDivisione CellulareGianmarco VindigniNessuna valutazione finora
- Cellule Procariote e Cellule EucarioteDocumento1 paginaCellule Procariote e Cellule EucarioteGianmarco VindigniNessuna valutazione finora
- Virus e PandemieDocumento8 pagineVirus e PandemieGianmarco VindigniNessuna valutazione finora
- Trascrizione DNA Traduzione RNA MutazioniDocumento7 pagineTrascrizione DNA Traduzione RNA MutazioniGianmarco VindigniNessuna valutazione finora
- Catalogo Guias Ttes. BettDocumento18 pagineCatalogo Guias Ttes. BettMarcelo ResckNessuna valutazione finora
- 1975 Fermentazioni AlcolicheDocumento37 pagine1975 Fermentazioni AlcolicheLucia LisciNessuna valutazione finora
- Manuale Tecnico Macchinisti Impianti A Fune.1686834618Documento148 pagineManuale Tecnico Macchinisti Impianti A Fune.1686834618Agnichandra SubediNessuna valutazione finora
- Autorizzazione Offline Di Ableton LiveDocumento1 paginaAutorizzazione Offline Di Ableton LiveFrancesco Groovisionary CapuaniNessuna valutazione finora
- 04 Comportamento FondamentiDocumento7 pagine04 Comportamento FondamentiingssNessuna valutazione finora
- Lezione07 DivideImpera RicercaBinaria MergeSortDocumento56 pagineLezione07 DivideImpera RicercaBinaria MergeSortsmegmusNessuna valutazione finora