Potrebbero piacerti anche
- Marchetti M., Pillastrini P. Neurofisiologia Del MovimentoDocumento550 pagineMarchetti M., Pillastrini P. Neurofisiologia Del MovimentoAttilio LiberatoNessuna valutazione finora
- Sport e scienze sociali: Fenomeni sportivi tra consumi, media e processi globaliDa EverandSport e scienze sociali: Fenomeni sportivi tra consumi, media e processi globaliNessuna valutazione finora
- 1° Corso - Osteopatia Craniale Vol 1 - Le Disfunzioni Della PDFDocumento89 pagine1° Corso - Osteopatia Craniale Vol 1 - Le Disfunzioni Della PDFpaolo68audisio100% (3)
- Disordini Del MovimentoDocumento138 pagineDisordini Del MovimentoorsolaNessuna valutazione finora
- Corpo e Movimento: La Consapevolezza Della Cultura Attraverso L'abito.Documento124 pagineCorpo e Movimento: La Consapevolezza Della Cultura Attraverso L'abito.MartinaPadrin0% (1)
- Contratti CompletoDocumento192 pagineContratti CompletosangambinoNessuna valutazione finora
- Piede e PodoscopioDocumento114 paginePiede e Podoscopiopaolo68audisioNessuna valutazione finora
- Corso Di Biochimica Generale - 13 Maggio 2012Documento199 pagineCorso Di Biochimica Generale - 13 Maggio 2012Irene SchenoneNessuna valutazione finora
- Il Sonno e Le Patologie Respiratorie PDFDocumento99 pagineIl Sonno e Le Patologie Respiratorie PDFPaoly PalmaNessuna valutazione finora
- CoronaVirus CoViD-19 SARS-CoV2 e Attivita Fisica Adattata: Considerazioni osservazioni curiosità scienza durante il periodo di quarantena di contenimento sociale della pandemiaDa EverandCoronaVirus CoViD-19 SARS-CoV2 e Attivita Fisica Adattata: Considerazioni osservazioni curiosità scienza durante il periodo di quarantena di contenimento sociale della pandemiaNessuna valutazione finora
- COMPLETO Gastroenterologia e Chirurgia Dell'Apparato DigerenteDocumento339 pagineCOMPLETO Gastroenterologia e Chirurgia Dell'Apparato Digerentebakshi88Nessuna valutazione finora
- Curs MCurs Mario Castagnini - Educare Neuromotorieario Castagnini - Educare NeuromotorieDocumento138 pagineCurs MCurs Mario Castagnini - Educare Neuromotorieario Castagnini - Educare NeuromotorieBiborka GyepesiNessuna valutazione finora
- Neuroscienze - Per Iniziare - Testo ComplementareDocumento125 pagineNeuroscienze - Per Iniziare - Testo ComplementareDavide MottaNessuna valutazione finora
- Necrosi e Autofagia CelulareDocumento86 pagineNecrosi e Autofagia Celularezolta9Nessuna valutazione finora
- Dal Big Bang A DioDocumento189 pagineDal Big Bang A DioMarcvs'9000% (1)
- Propriocezione e Movimento (Appunti Del 2003, 14 Pagine) PDFDocumento14 paginePropriocezione e Movimento (Appunti Del 2003, 14 Pagine) PDFrosaNessuna valutazione finora
- 14 InvecchiamentoDocumento33 pagine14 Invecchiamentobakshi88100% (1)
- FaringeDocumento5 pagineFaringebakshi88Nessuna valutazione finora
- La Cellula - WikibookDocumento107 pagineLa Cellula - WikibookMichele Scipioni100% (1)
- VirologiaDocumento151 pagineVirologiaSimoiadeNessuna valutazione finora
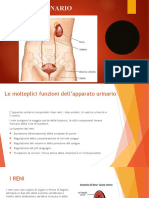
- Apparato UrinarioDocumento17 pagineApparato UrinarioCate?Nessuna valutazione finora
- La Rieducazione Dopo Intervento Di Ernia DiscaleDocumento13 pagineLa Rieducazione Dopo Intervento Di Ernia DiscalePaolo TintiNessuna valutazione finora
- La Grande ViaDocumento8 pagineLa Grande ViaLucia PenazzoNessuna valutazione finora
- Farmacologia ApplicataDocumento151 pagineFarmacologia ApplicataMukkamilkNessuna valutazione finora
- Trovare Su Internet - Fabio BrivioDocumento127 pagineTrovare Su Internet - Fabio BrivioSalvatore CaciNessuna valutazione finora
- Fisiologia DomandeDocumento17 pagineFisiologia DomandeLuca BevarNessuna valutazione finora
- Ernia InguinaleDocumento70 pagineErnia InguinaleArduino RocciolettiNessuna valutazione finora
- DiabeteDocumento74 pagineDiabeteapi-3863584Nessuna valutazione finora
- Canale InguinaleDocumento54 pagineCanale InguinaleKreshnik Idrizaj100% (1)
- Elementi Di Mobilità Articolare e Di Elasticità Muscolare 17 Dicembre 23 00Documento30 pagineElementi Di Mobilità Articolare e Di Elasticità Muscolare 17 Dicembre 23 00gabriel ungureanuNessuna valutazione finora
- Proudhon L'idea Generale Di Rivoluzione Nel XIX SecoloDocumento260 pagineProudhon L'idea Generale Di Rivoluzione Nel XIX SecoloWalter SimonettiNessuna valutazione finora
- Sintesi Malattie Del SangueDocumento8 pagineSintesi Malattie Del SangueemypippoNessuna valutazione finora
- Artride Reumatoide RiabilitazioneDocumento48 pagineArtride Reumatoide RiabilitazioneGiuseppe Foggia100% (1)
- Reflusso Gastro-Esofageo Tratt KinesiterapicoDocumento6 pagineReflusso Gastro-Esofageo Tratt KinesiterapicoRoberto BernyNessuna valutazione finora
- Apparato Locomotore PDFDocumento16 pagineApparato Locomotore PDFginobarbarohotmail.comNessuna valutazione finora
- Società Italiana Paraplegia - Gli Indicatori Di OutcomeDocumento25 pagineSocietà Italiana Paraplegia - Gli Indicatori Di OutcomeBruna Ortiz100% (1)
- Energia e Fissione NucleareDocumento105 pagineEnergia e Fissione NucleareAdailton ManfiesNessuna valutazione finora
- Appunti RachideDocumento6 pagineAppunti RachideMaria Serena RanaNessuna valutazione finora
- Geologia 2Documento115 pagineGeologia 2Allegra DelmonteNessuna valutazione finora
- Sistema Gastrointestinale Fisiologia DelDocumento160 pagineSistema Gastrointestinale Fisiologia DelMiriam GalliNessuna valutazione finora
- Introduzione TEM 2021Documento15 pagineIntroduzione TEM 2021TomyasuNessuna valutazione finora
- Scheda Tecnica n.21.1Documento2 pagineScheda Tecnica n.21.1Loredana SansaviniNessuna valutazione finora
- Biochimica ClinicaDocumento20 pagineBiochimica ClinicaUlvionaNessuna valutazione finora
- Tesi F. D'EliaDocumento143 pagineTesi F. D'Eliayeast60Nessuna valutazione finora
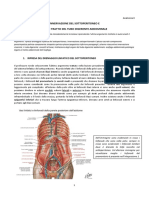
- 39-Anatomia II-06.04.2016-Innervazione Del Sottoperitoneo e Trattazione Del Tubo Digerente AddominaleDocumento13 pagine39-Anatomia II-06.04.2016-Innervazione Del Sottoperitoneo e Trattazione Del Tubo Digerente AddominaleSandroNessuna valutazione finora
- Rge DiapositiveDocumento40 pagineRge DiapositiveValentina FallarinoNessuna valutazione finora
- Biologia Applicata - Biologia Dello SviluppoDocumento59 pagineBiologia Applicata - Biologia Dello Sviluppoapi-3826003100% (4)
- Fisiopatologia Del Sistema MotorioDocumento58 pagineFisiopatologia Del Sistema MotorioIlaria PuttiniNessuna valutazione finora
- Fondamenti Di Nutrizione UmanaDocumento10 pagineFondamenti Di Nutrizione Umanarufus991Nessuna valutazione finora
- Ittero OstruttivoDocumento43 pagineIttero Ostruttivoapi-3733154100% (1)
- Fisiopatologia Chirurgica Dell' EsofagoDocumento14 pagineFisiopatologia Chirurgica Dell' EsofagoBarbara MelchiorettoNessuna valutazione finora
- Dispensa - Fisiologia Dell'esercizio Muscolare PDFDocumento66 pagineDispensa - Fisiologia Dell'esercizio Muscolare PDFFrancesco MemoliNessuna valutazione finora
- Appunti Sul Cibo Di Andrea BraggioDocumento17 pagineAppunti Sul Cibo Di Andrea BraggioandreabraggioNessuna valutazione finora
- Midollo Spinale - Vie Ascendenti e DiscendentiDocumento2 pagineMidollo Spinale - Vie Ascendenti e DiscendentiDaniele VintiNessuna valutazione finora
- Trasformazioni Del Corpo in GravidanzaDocumento18 pagineTrasformazioni Del Corpo in GravidanzaLaura Fearsome ViewNessuna valutazione finora
- Sistema VestibolareDocumento31 pagineSistema Vestibolaresimo_pNessuna valutazione finora
- Guida PhotofiltreDocumento7 pagineGuida PhotofiltrefukinagashiNessuna valutazione finora
- Piede PiattoDocumento15 paginePiede PiattoHermannNessuna valutazione finora
- PNEUMOLOGIADocumento43 paginePNEUMOLOGIAFedericoPppzPippoNessuna valutazione finora
- Ernia DiscaleDocumento16 pagineErnia Discalealfredo81itNessuna valutazione finora
- La Sindrome del Colon Irritabile: Guida alla fisiopatologia, alimentazione ed integrazioneDa EverandLa Sindrome del Colon Irritabile: Guida alla fisiopatologia, alimentazione ed integrazioneNessuna valutazione finora
- COMPLETO EmatologiaDocumento194 pagineCOMPLETO Ematologiabakshi88Nessuna valutazione finora
- MEDICINA Robbins - FegatoDocumento46 pagineMEDICINA Robbins - Fegatobakshi88Nessuna valutazione finora
- 20 - Tecniche BiomolecolariDocumento12 pagine20 - Tecniche Biomolecolaribakshi88Nessuna valutazione finora
- 19 - Tecniche BiochimicheDocumento13 pagine19 - Tecniche Biochimichebakshi88Nessuna valutazione finora
- Visual SystemDocumento164 pagineVisual Systembakshi88Nessuna valutazione finora
- FaringeDocumento5 pagineFaringebakshi88Nessuna valutazione finora
- 29 BisDocumento41 pagine29 Bissupernatura100% (5)
- Tabella Vie SNCDocumento3 pagineTabella Vie SNCLeonardo Pzl100% (1)
- Lezione 1 NeuroradiologiaDocumento13 pagineLezione 1 NeuroradiologiaFraNessuna valutazione finora
- Le 5 Leggi Di Hamer-Medicina AlternativaDocumento4 pagineLe 5 Leggi Di Hamer-Medicina AlternativaVico Massagista e Quiropraxia - Massagem, Massoterapia e Ventosa Terapia - São José SC.100% (1)
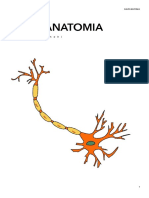
- NEUROANATOMIADocumento172 pagineNEUROANATOMIAFrancesco LanzaNessuna valutazione finora
- Sistema Nervoso RiassuntoDocumento64 pagineSistema Nervoso Riassuntolucchini.paolo3002Nessuna valutazione finora
- Anatomia II Medicina PisaDocumento100 pagineAnatomia II Medicina PisanuunumclNessuna valutazione finora
- CircolatorioDocumento41 pagineCircolatorioRiccardo Mason TroiaNessuna valutazione finora
- NeuroDocumento9 pagineNeuroMatteo FerrariNessuna valutazione finora
- Psicologia MusicaleDocumento5 paginePsicologia MusicaleAttilio PerfettiNessuna valutazione finora
- Midollo TroncoDocumento15 pagineMidollo Troncoeve_612Nessuna valutazione finora
- Sistema Nervoso Encefalo e Nervi Cranici Capitolo 15Documento27 pagineSistema Nervoso Encefalo e Nervi Cranici Capitolo 15enrico73Nessuna valutazione finora
- Anatomia Del Sistema Nervoso Centrale - Schemi VisiviDocumento58 pagineAnatomia Del Sistema Nervoso Centrale - Schemi VisiviClara ScopellitiNessuna valutazione finora
- GRAFOLOGIADocumento4 pagineGRAFOLOGIAJoePansaNessuna valutazione finora
- Lezione 03 - Neurologia - 2021-2022 Semeiologia Funzione Motoria Gangli Della Base Coordinazione. Funzione SensitivaDocumento124 pagineLezione 03 - Neurologia - 2021-2022 Semeiologia Funzione Motoria Gangli Della Base Coordinazione. Funzione Sensitivamaria cristina ristagnoNessuna valutazione finora
- La Biologia Delle Emozioni 9788896865422 637111Documento225 pagineLa Biologia Delle Emozioni 9788896865422 637111Elisa Osimani100% (4)
- TTD - TMU - Lezioni I - XIIDocumento81 pagineTTD - TMU - Lezioni I - XIIMaegazine EnizageamNessuna valutazione finora
- Neoplasie CerebraliDocumento12 pagineNeoplasie CerebraliPatrizia CastellanetaNessuna valutazione finora
- Grandi Vie NervoseDocumento15 pagineGrandi Vie NervoseAnonymous 9aq7eNvONessuna valutazione finora
- Appunti NeruopsicologiaDocumento32 pagineAppunti NeruopsicologiaMartina CalvoNessuna valutazione finora
- FisiologiaDocumento4 pagineFisiologiaHghh NgthhNessuna valutazione finora