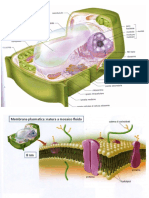
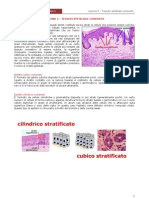
Potrebbero piacerti anche
- Morte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cDa EverandMorte cellulare programmata indotta da acido acetico in Saccharomyces cerevisiae: produzione di ROS e rilascio di citocromo cNessuna valutazione finora
- La Cellula VegetaleDocumento47 pagineLa Cellula VegetaleEleonora AngelocolaNessuna valutazione finora
- 1 La Cellula VegetaleDocumento3 pagine1 La Cellula VegetaleAsia ZaccariaNessuna valutazione finora
- Biologia 27 - Membrana Cellulare - AdesioneDocumento3 pagineBiologia 27 - Membrana Cellulare - AdesioneFrancesco TarulloNessuna valutazione finora
- La Parete Cellulare-Prima ParteDocumento40 pagineLa Parete Cellulare-Prima PartenomeeeeNessuna valutazione finora
- BotanicaDocumento40 pagineBotanicaArturo FiorenzaNessuna valutazione finora
- Il Tessuto EpitelialeDocumento6 pagineIl Tessuto EpitelialeDiletta De lucaNessuna valutazione finora
- TESSUTIDocumento15 pagineTESSUTIRosa NoceraNessuna valutazione finora
- 2 I TessutiDocumento3 pagine2 I TessutiAsia ZaccariaNessuna valutazione finora
- Schemi Anatomia ComparataDocumento42 pagineSchemi Anatomia ComparataEleonora CrespiaticoNessuna valutazione finora
- La Matrice EXTRACELLULAREDocumento5 pagineLa Matrice EXTRACELLULAREManuel RodriguezNessuna valutazione finora
- Biologia VegataleDocumento9 pagineBiologia VegataleiwttbaNessuna valutazione finora
- Le Cellule Eucariotiche - BookinprogressDocumento12 pagineLe Cellule Eucariotiche - BookinprogressCarmenNessuna valutazione finora
- I Tessuti ConnettiviDocumento5 pagineI Tessuti ConnettiviLauraNessuna valutazione finora
- Istologia Appunti - OdtDocumento20 pagineIstologia Appunti - Odtprova123100% (1)
- La CellulaDocumento10 pagineLa CellulaAngelica CoppolaNessuna valutazione finora
- 2 - Sostegno - 04-4-19Documento51 pagine2 - Sostegno - 04-4-19tomaoana011Nessuna valutazione finora
- Differenziamento Cellulare - Derivazione Embrionale IstologiaDocumento17 pagineDifferenziamento Cellulare - Derivazione Embrionale IstologiaWicked_KnightNessuna valutazione finora
- CELLULADocumento5 pagineCELLULAStanislav VovkNessuna valutazione finora
- Tejidos GeneserDocumento27 pagineTejidos GeneserHittler ZahiDNessuna valutazione finora
- ScienzeDocumento9 pagineScienzeBeatrice SanavioNessuna valutazione finora
- Lezione 2 - Tessuto Epiteliale CompostoDocumento7 pagineLezione 2 - Tessuto Epiteliale CompostotrillicosaNessuna valutazione finora
- La CellulaDocumento27 pagineLa CellulaAlessio PasquaNessuna valutazione finora
- Citologia e Istologia RiassuntoDocumento45 pagineCitologia e Istologia RiassuntoChiara De MartinoNessuna valutazione finora
- 1) La Cellula Vegetale FVDocumento51 pagine1) La Cellula Vegetale FVapi-26155364100% (2)
- Biologia - Riassunti Di Karp, "Biologia Cellulare e Molecolare"Documento75 pagineBiologia - Riassunti Di Karp, "Biologia Cellulare e Molecolare"Matteo Paolucci100% (3)
- Biologia CellulareDocumento5 pagineBiologia CellulareMarzia ZucchelliNessuna valutazione finora
- BiotecnologieDocumento55 pagineBiotecnologieporfilo68Nessuna valutazione finora
- IstologiaDocumento109 pagineIstologiaporfilo68Nessuna valutazione finora
- MitosiDocumento66 pagineMitosiporfilo68Nessuna valutazione finora
- InsulinaDocumento55 pagineInsulinaporfilo68Nessuna valutazione finora
- FecondazioneDocumento92 pagineFecondazioneporfilo68Nessuna valutazione finora
- Ciclo OvaricoDocumento94 pagineCiclo Ovaricoporfilo68Nessuna valutazione finora
- Parete Cellulare BVF Ott 20Documento63 pagineParete Cellulare BVF Ott 20Eleonora AngelocolaNessuna valutazione finora
- Appunti BiochimicaDocumento2 pagineAppunti Biochimicafrancescagarelli00Nessuna valutazione finora
- Biologia Lezione 5Documento9 pagineBiologia Lezione 5alessandro.massar134Nessuna valutazione finora
- MICROBIOLOGIA - Membrana e Parete BattericaDocumento5 pagineMICROBIOLOGIA - Membrana e Parete BattericaOmmalleyNessuna valutazione finora
- Tessuto CartilagineoDocumento3 pagineTessuto CartilagineoDiletta De lucaNessuna valutazione finora
- I Tessuti VegetaliDocumento13 pagineI Tessuti Vegetalialice9980Nessuna valutazione finora
- UntitledDocumento51 pagineUntitledSofia AmentaNessuna valutazione finora
- Tessuto ConnettivoDocumento4 pagineTessuto Connettivomarta nespoliNessuna valutazione finora
- CitologiaDocumento6 pagineCitologiajobronyNessuna valutazione finora
- 9 Eucariote Animale - Citoscheletro, CentrioliDocumento8 pagine9 Eucariote Animale - Citoscheletro, CentrioliLuana LeonardiNessuna valutazione finora
- Eucarioti, Procarioti e Virus PDFDocumento12 pagineEucarioti, Procarioti e Virus PDFGuido CaligioreNessuna valutazione finora
- 01 La CellulaDocumento7 pagine01 La CellulaAlessio LampisNessuna valutazione finora
- Citoscheletro: Miosina II È Responsabile Della Contrazione e Della Citodieresi. Le Miosine I e V Sono Responsabili DelDocumento14 pagineCitoscheletro: Miosina II È Responsabile Della Contrazione e Della Citodieresi. Le Miosine I e V Sono Responsabili DelRosi De LucaNessuna valutazione finora
- Copia Di La CellulaDocumento6 pagineCopia Di La CelluladomenicaNessuna valutazione finora
- Appunti BiologiaDocumento12 pagineAppunti BiologiaAlessia PolitoNessuna valutazione finora
- File Completo IstologiaDocumento56 pagineFile Completo IstologiaSara Chiavacci100% (1)
- Citologia-Generalità e MembranaDocumento32 pagineCitologia-Generalità e MembranaHamidreza DeldadehNessuna valutazione finora
- Capitolo 1. L Invecchiamento Della Retina. Introduzione. Organizzazione Morfologica e Funzionale Della Retina PDFDocumento161 pagineCapitolo 1. L Invecchiamento Della Retina. Introduzione. Organizzazione Morfologica e Funzionale Della Retina PDFtopobucciaNessuna valutazione finora
- TessutiDocumento64 pagineTessutiDestroyer 99Nessuna valutazione finora
- Connettivi Di IstologiaDocumento19 pagineConnettivi Di Istologiacinthia8989Nessuna valutazione finora
- Istologia - Schema RiassuntivoDocumento19 pagineIstologia - Schema Riassuntivoveruska_93100% (2)
- Dominio Basale e Le Sue SpecializzazioniDocumento1 paginaDominio Basale e Le Sue SpecializzazioniAlib GrantNessuna valutazione finora
- Istologia Ps Classi 2,3,4 2019-2020Documento304 pagineIstologia Ps Classi 2,3,4 2019-2020Matteo RussoNessuna valutazione finora
- Citologia e HistologiaDocumento36 pagineCitologia e Histologiasthefanycp19Nessuna valutazione finora
- L’alcaloide vegetale Voacamina esercita un effetto chemiosensibilizzante su cellule tumorali farmacoresistenti mediante l’induzione del processo autofagicoDa EverandL’alcaloide vegetale Voacamina esercita un effetto chemiosensibilizzante su cellule tumorali farmacoresistenti mediante l’induzione del processo autofagicoNessuna valutazione finora
- Ringiovanire Con Il Plasma Sanguigno Dei GiovaniDa EverandRingiovanire Con Il Plasma Sanguigno Dei GiovaniNessuna valutazione finora
- IstologiaDocumento47 pagineIstologiaSimone Chinaglia0% (1)
- Lezione 4Documento34 pagineLezione 4api-26155364100% (1)
- Appunti Citologia e IstologiaDocumento73 pagineAppunti Citologia e IstologiaAntonio LambresaNessuna valutazione finora
- Microbiologia - Fac. BiotecnologieDocumento39 pagineMicrobiologia - Fac. BiotecnologieAgnese SanninoNessuna valutazione finora