Potrebbero piacerti anche
- La seconda prova di matematica e fisica: svolgimento commentato delle prove ufficiali 2019Da EverandLa seconda prova di matematica e fisica: svolgimento commentato delle prove ufficiali 2019Nessuna valutazione finora
- 13-Il-Test-A-Tre-Punti Genetica Agraria UnissDocumento16 pagine13-Il-Test-A-Tre-Punti Genetica Agraria UnissAlessandra Patricia LopesNessuna valutazione finora
- Calcolo Distanze Di Mappa Incrocio A Tre Punti InterferenzaDocumento7 pagineCalcolo Distanze Di Mappa Incrocio A Tre Punti InterferenzaTonyNessuna valutazione finora
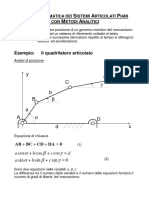
- Teorema Di Carnot Nella Risoluzione Dei TriangoliDocumento8 pagineTeorema Di Carnot Nella Risoluzione Dei TriangoliClaudi SeeyyeNessuna valutazione finora
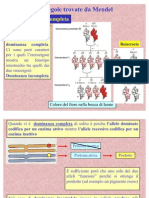
- 2 Mendel Con EserciziDocumento17 pagine2 Mendel Con EserciziMichela PintoNessuna valutazione finora
- Teorema Dei Seni e Di CarnotDocumento1 paginaTeorema Dei Seni e Di CarnotSebastiano CaltabianoNessuna valutazione finora
- Risoluzione Dei TriangoliDocumento3 pagineRisoluzione Dei TriangolikimNessuna valutazione finora
- 01 DielettriciDocumento4 pagine01 DielettriciRobertoNessuna valutazione finora
- Trigo MetriDocumento43 pagineTrigo MetriDedi ResoNessuna valutazione finora
- 2015 LuglioDocumento4 pagine2015 LuglioSara MounaddimeNessuna valutazione finora
- Ponte Di WheatstoneDocumento22 paginePonte Di WheatstoneLuigi ReveruzziNessuna valutazione finora
- Quiz BiotecnologieDocumento6 pagineQuiz BiotecnologieSam VenNessuna valutazione finora
- Mate 1classe 30es ZanichelliDocumento7 pagineMate 1classe 30es ZanichelliMarco FaellaNessuna valutazione finora
- Elce-Teorema Seni Teorema CosenoDocumento10 pagineElce-Teorema Seni Teorema CosenoVincenzo FerraraNessuna valutazione finora
- Mappatura GeneticaDocumento60 pagineMappatura GeneticafarloccoloccoNessuna valutazione finora
- 4B FIS Teoria BDocumento2 pagine4B FIS Teoria BAlessioRocciNessuna valutazione finora
- Dispensa PDFDocumento18 pagineDispensa PDFDestinyNessuna valutazione finora
- S Successioni 0405 Q StraDocumento2 pagineS Successioni 0405 Q Strajarevir820Nessuna valutazione finora
- Ponte Di ThompsonDocumento3 paginePonte Di ThompsonDanilo InfantinoNessuna valutazione finora
- M01a - Sintesi Planetario 1Documento2 pagineM01a - Sintesi Planetario 1perty92Nessuna valutazione finora
- Ver 4 4 DicDocumento2 pagineVer 4 4 DicGix Fredck MaliNessuna valutazione finora
- Lez 49Documento7 pagineLez 49Francesco PreteNessuna valutazione finora
- Esempio Test LaibDocumento4 pagineEsempio Test LaibAndrea SpitaleNessuna valutazione finora
- Trigonometria 1Documento18 pagineTrigonometria 1CarlosCoñacaQuispeNessuna valutazione finora
- Balotaio 2022-IIDocumento61 pagineBalotaio 2022-IIWilson Mamani CondoriNessuna valutazione finora
- Prova 1: Sei Pronto Per La Verifica? - Rette Perpendicolari e Rette Parallele - Riepilogo (Pag. G137)Documento3 pagineProva 1: Sei Pronto Per La Verifica? - Rette Perpendicolari e Rette Parallele - Riepilogo (Pag. G137)Alessandro PettiNessuna valutazione finora
- Misureb Lezione Circuito RCDocumento21 pagineMisureb Lezione Circuito RCsalvo castiglioneNessuna valutazione finora
- Esercitazione Prove D'esameDocumento10 pagineEsercitazione Prove D'esameMatheus PadilhaNessuna valutazione finora
- Fondamenti Di Meccanica e Biomeccanica Del Professore Leban - Lez 6 - Aa 2018-19Documento11 pagineFondamenti Di Meccanica e Biomeccanica Del Professore Leban - Lez 6 - Aa 2018-19antonioNessuna valutazione finora
- FormularioDocumento8 pagineFormularioAllegra PirasNessuna valutazione finora
- 4B FIS Teoria ADocumento2 pagine4B FIS Teoria AAlessioRocciNessuna valutazione finora
- Spianamenti OrizzontaliDocumento11 pagineSpianamenti OrizzontaliLucianoNessuna valutazione finora
- Sensori Al QuarzoDocumento19 pagineSensori Al QuarzoFrancoNessuna valutazione finora
- Archimede 2023 TriennioDocumento2 pagineArchimede 2023 TrienniototogualanoNessuna valutazione finora
- Formulario Di Analisi MatematicaDocumento95 pagineFormulario Di Analisi MatematicaElisNessuna valutazione finora
- Risoluzione Dei Triangoli QualsiasiDocumento2 pagineRisoluzione Dei Triangoli QualsiasiValeria La RoccaNessuna valutazione finora
- Verifica Cerchio 1-3Documento3 pagineVerifica Cerchio 1-3Debora Panzarella100% (1)
- Cap8 Esercizi RisoltiDocumento32 pagineCap8 Esercizi Risoltiandrea_luca_51Nessuna valutazione finora
- Prova 1: Sei Pronto Per La Verifica? - Triangoli - 2 (Pag. G89)Documento5 pagineProva 1: Sei Pronto Per La Verifica? - Triangoli - 2 (Pag. G89)Alessandro PettiNessuna valutazione finora
- Verifica Dicembre MonomiDocumento1 paginaVerifica Dicembre MonomiYusef ElshourbagiNessuna valutazione finora
- Test - Fisica II - 2013-2014 - Prima - ParteDocumento28 pagineTest - Fisica II - 2013-2014 - Prima - ParteajejeNessuna valutazione finora
- Stem 08 07 2021 A028 SardegnaDocumento27 pagineStem 08 07 2021 A028 SardegnaNicola CoppolinoNessuna valutazione finora
- Formulario: Trigonometria. Risoluzione Dei Triangoli: Math - ItDocumento2 pagineFormulario: Trigonometria. Risoluzione Dei Triangoli: Math - ItfaNessuna valutazione finora
- MATEMATICADocumento10 pagineMATEMATICAriccardo.bindiNessuna valutazione finora
- Test Economia Ferrara 2012 2013Documento4 pagineTest Economia Ferrara 2012 2013Tommaso CatellaniNessuna valutazione finora
- Bignami AlgebraDocumento17 pagineBignami AlgebraFrancesca De PariNessuna valutazione finora
- Test Fisica II 2014-2015Documento72 pagineTest Fisica II 2014-2015Raffaele PastoreNessuna valutazione finora
- Albero Iperstatico 2Documento8 pagineAlbero Iperstatico 2emmene freNessuna valutazione finora
- Prova 1: Sei Pronto Per La Verifica? - Triangoli - Riepilogo (Pag. G95)Documento5 pagineProva 1: Sei Pronto Per La Verifica? - Triangoli - Riepilogo (Pag. G95)Alessandro Petti100% (1)
- Esercitazione 5 PDFDocumento1 paginaEsercitazione 5 PDFdomNessuna valutazione finora
- Teoremi Triangoli QualsiasiDocumento1 paginaTeoremi Triangoli QualsiasiMartina FassoNessuna valutazione finora
- Esercizi Risolti Prova Scritta 2019 2 21Documento4 pagineEsercizi Risolti Prova Scritta 2019 2 21Michela FioreNessuna valutazione finora
- Determinanti TrigonometriciDocumento5 pagineDeterminanti TrigonometriciMadalina Mihaela GalavanNessuna valutazione finora
- 22SQ Bell PDFDocumento17 pagine22SQ Bell PDFSono ioNessuna valutazione finora
- La Circonferenza Nel Piano CartesianoDocumento34 pagineLa Circonferenza Nel Piano CartesianorosagiuliaNessuna valutazione finora
- Esercizi Fisica 2Documento4 pagineEsercizi Fisica 2FilippoNessuna valutazione finora
- PoliquizDocumento13 paginePoliquizajejeNessuna valutazione finora
- Dal - Fenotipo - Al - Genotipo 2Documento6 pagineDal - Fenotipo - Al - Genotipo 2Georgiana ReolonNessuna valutazione finora
- Test ImpiantiDocumento5 pagineTest ImpiantiMarco MuratoreNessuna valutazione finora
- Archimede 2023 BiennioDocumento2 pagineArchimede 2023 BienniobehnashamsNessuna valutazione finora
- Metodologia Medicina Di LaboratorioDocumento25 pagineMetodologia Medicina Di LaboratoriofarloccoloccoNessuna valutazione finora
- Approccio PrenataleDocumento34 pagineApproccio PrenatalefarloccoloccoNessuna valutazione finora
- Lez.3 Biochimica ClinicaDocumento12 pagineLez.3 Biochimica ClinicafarloccoloccoNessuna valutazione finora
- Prob Bayes Test!Documento49 pagineProb Bayes Test!farloccoloccoNessuna valutazione finora
- Quiz SSBCDocumento100 pagineQuiz SSBCfarloccoloccoNessuna valutazione finora
- Duchenne Sma e TerapieDocumento74 pagineDuchenne Sma e TerapiefarloccoloccoNessuna valutazione finora
- Mappatura GeneticaDocumento60 pagineMappatura GeneticafarloccoloccoNessuna valutazione finora
- Quiz Patologia Clinica 2Documento108 pagineQuiz Patologia Clinica 2farloccoloccoNessuna valutazione finora
- Tracce Ispettore-Tecnico PDFDocumento14 pagineTracce Ispettore-Tecnico PDFLuis PepsNessuna valutazione finora
- Polizia ScientificaDocumento10 paginePolizia ScientificafarloccoloccoNessuna valutazione finora
- Direttore Tecnico Biologo TemiDocumento1 paginaDirettore Tecnico Biologo TemifarloccoloccoNessuna valutazione finora
- Nome CognomeDocumento1 paginaNome CognomefarloccoloccoNessuna valutazione finora
- Tracce BiologiDocumento7 pagineTracce BiologifarloccoloccoNessuna valutazione finora
- HaccpDocumento5 pagineHaccpfarloccoloccoNessuna valutazione finora
- Cosa Studiare e Dove StudiareDocumento2 pagineCosa Studiare e Dove StudiarefarloccoloccoNessuna valutazione finora
- 12 Mitosi Meiosi-UniPDDocumento43 pagine12 Mitosi Meiosi-UniPDDonato PaglionicoNessuna valutazione finora
- Glossario Biologia Zanichelli Online Per La Scuola CompressDocumento5 pagineGlossario Biologia Zanichelli Online Per La Scuola CompressRosaria AlbaNessuna valutazione finora
- Quaderno Biologia Scuole SuperioriDocumento25 pagineQuaderno Biologia Scuole SuperioriFabian TomasiNessuna valutazione finora
- Mitosi e Meiosi - Dispensa Di BiologiaDocumento1 paginaMitosi e Meiosi - Dispensa Di BiologiaSimone SecondoNessuna valutazione finora
- Genetica 1Documento175 pagineGenetica 1Luca BaroniNessuna valutazione finora
- PancaldiDocumento5 paginePancaldipierucci93Nessuna valutazione finora
- Linkage GeneticaDocumento32 pagineLinkage GeneticaJessica PopeNessuna valutazione finora
- QuizfarmaDocumento25 pagineQuizfarmaSam VenNessuna valutazione finora
- Genetic ADocumento101 pagineGenetic AterezaNessuna valutazione finora
- Biologia e Genetica AppuntiDocumento25 pagineBiologia e Genetica AppuntiJessica Talpo100% (1)
- Concetti Di Genetica MedicaDocumento28 pagineConcetti Di Genetica Medicajuri84Nessuna valutazione finora
- Biologia 17 - Genetica Classica PDFDocumento6 pagineBiologia 17 - Genetica Classica PDFkostymassNessuna valutazione finora
- 12 Esperimento Di Bateson e Punnet e Associazione Genetica Genetica Agraria UnissDocumento15 pagine12 Esperimento Di Bateson e Punnet e Associazione Genetica Genetica Agraria UnissAlessandra Patricia LopesNessuna valutazione finora
- MeiosiDocumento40 pagineMeiosiGiuseppeNessuna valutazione finora
- Genetica - Livello AvanzatoDocumento47 pagineGenetica - Livello AvanzatokostymassNessuna valutazione finora
- EserciziDocumento46 pagineEserciziLaura RossiNessuna valutazione finora
- Genetica Del CanarinoDocumento22 pagineGenetica Del Canarinoghino_di_tacco100% (1)
- Biologia Prof. RinaldiDocumento21 pagineBiologia Prof. RinaldiJonella D'agataNessuna valutazione finora
- BIOLOGIA - Biologia - Genetica (Mendel)Documento25 pagineBIOLOGIA - Biologia - Genetica (Mendel)cezaol4428Nessuna valutazione finora
- Lezione03sn PDFDocumento33 pagineLezione03sn PDFterezaNessuna valutazione finora
- Biotech Cellulari, Molecolari e Computazionali - PaolellaDocumento46 pagineBiotech Cellulari, Molecolari e Computazionali - PaolellaAntonio Cox FerraroNessuna valutazione finora
- Genetica MedicaDocumento151 pagineGenetica MedicaBeatrice CarliniNessuna valutazione finora
- Le Tappe Della Meiosi: Divisione Equazionale, Simile Alla MiDocumento3 pagineLe Tappe Della Meiosi: Divisione Equazionale, Simile Alla MiPierluigi CesarioNessuna valutazione finora
- Meiosi e MitosiDocumento3 pagineMeiosi e MitosielixdxNessuna valutazione finora
- Riassunto Embriologia Umana PDFDocumento51 pagineRiassunto Embriologia Umana PDFEugenia DantonioNessuna valutazione finora
- C12 MeiosiDocumento38 pagineC12 MeiosiAnonymous VoYfGuoNessuna valutazione finora
- Dispensa GeneticaDocumento185 pagineDispensa GeneticaDANIELA TOMASELLINessuna valutazione finora
- Nomofobia: Cos'è, chi ne soffre, perché esisteDa EverandNomofobia: Cos'è, chi ne soffre, perché esisteNessuna valutazione finora
- Comprendere i messaggi delle nostre articolazioni: Prevenire e curaretensioni e doloriscoprendo il legametra giunture, emozionie comportamentiDa EverandComprendere i messaggi delle nostre articolazioni: Prevenire e curaretensioni e doloriscoprendo il legametra giunture, emozionie comportamentiNessuna valutazione finora
- Agnihotri - I Custodi del Fuoco: Il Sentiero di Guarigione del Guerriero del FuocoDa EverandAgnihotri - I Custodi del Fuoco: Il Sentiero di Guarigione del Guerriero del FuocoNessuna valutazione finora
- Come essere felici in un mondo di merda: Manuale di sopravvivenza spirituale nel mondo modernoDa EverandCome essere felici in un mondo di merda: Manuale di sopravvivenza spirituale nel mondo modernoNessuna valutazione finora
- Biologia quantica: Viaggio ai confini della guarigioneDa EverandBiologia quantica: Viaggio ai confini della guarigioneNessuna valutazione finora
- The Secret of Spirit-Thought Magic - Now-Project the thoughts that summon spirits from the Magic Astral World to make your secret wishes come true!Da EverandThe Secret of Spirit-Thought Magic - Now-Project the thoughts that summon spirits from the Magic Astral World to make your secret wishes come true!Valutazione: 5 su 5 stelle5/5 (6)
- La teoria della terra cava. The hollow earth theoryDa EverandLa teoria della terra cava. The hollow earth theoryValutazione: 3 su 5 stelle3/5 (1)
- Il trattamento fasciale osteopatico per il sistema nervoso centrale e perifericoDa EverandIl trattamento fasciale osteopatico per il sistema nervoso centrale e perifericoValutazione: 5 su 5 stelle5/5 (1)
- Epigenetica Per Principianti e Intermedi (2 Libri in 1). Come l'epigenetica può potenzialmente rivoluzionare la nostra comprensione della struttura e del comportamento della vita biologica sulla TerraDa EverandEpigenetica Per Principianti e Intermedi (2 Libri in 1). Come l'epigenetica può potenzialmente rivoluzionare la nostra comprensione della struttura e del comportamento della vita biologica sulla TerraNessuna valutazione finora
- Il miracolo della medicina rigenerativa: Come invertire il processo d’invecchiamento in modo naturaleDa EverandIl miracolo della medicina rigenerativa: Come invertire il processo d’invecchiamento in modo naturaleNessuna valutazione finora
- Come realizzare cose meravigliose … in un mondo di merda!: Metodi pratici per crescere, realizzare, cambiare le cose... ... anche quando tutto il mondo ci rema contro!Da EverandCome realizzare cose meravigliose … in un mondo di merda!: Metodi pratici per crescere, realizzare, cambiare le cose... ... anche quando tutto il mondo ci rema contro!Nessuna valutazione finora
- Tisane, Decotti e Infusi in terrazza: Curarsi con le tisaneDa EverandTisane, Decotti e Infusi in terrazza: Curarsi con le tisaneNessuna valutazione finora
- Risvegliando Gaia, La griglia del cristallo di Lemuria: La griglia del cristallo di LemuriaDa EverandRisvegliando Gaia, La griglia del cristallo di Lemuria: La griglia del cristallo di LemuriaNessuna valutazione finora
- Economia, ambiente e criminalità nel MezzogiornoDa EverandEconomia, ambiente e criminalità nel MezzogiornoValutazione: 5 su 5 stelle5/5 (1)
- Io comando, il mio fisico obbedisce e migliora: Alla ricerca di una vita longevaDa EverandIo comando, il mio fisico obbedisce e migliora: Alla ricerca di una vita longevaNessuna valutazione finora
- The Church of Darwin: Dogmas of evolution and the scientists that criticize itDa EverandThe Church of Darwin: Dogmas of evolution and the scientists that criticize itNessuna valutazione finora
- MARCO TODESCHINI - Tra Fisica e MetafisicaDa EverandMARCO TODESCHINI - Tra Fisica e MetafisicaValutazione: 5 su 5 stelle5/5 (1)
- Il Risveglio e il gioco cosmico dell’uomo: Cos’è il cuore cristallino della Terra?Da EverandIl Risveglio e il gioco cosmico dell’uomo: Cos’è il cuore cristallino della Terra?Nessuna valutazione finora
- Neuroscienze per Coach: Come usare le più recenti ricerche a beneficio dei tuoi clientiDa EverandNeuroscienze per Coach: Come usare le più recenti ricerche a beneficio dei tuoi clientiNessuna valutazione finora