Documenti di Didattica
Documenti di Professioni
Documenti di Cultura
Docsity Sbobine Funzione Renale Fisiologia
Caricato da
Enrico CusinoTitolo originale
Copyright
Formati disponibili
Condividi questo documento
Condividi o incorpora il documento
Hai trovato utile questo documento?
Questo contenuto è inappropriato?
Segnala questo documentoCopyright:
Formati disponibili
Docsity Sbobine Funzione Renale Fisiologia
Caricato da
Enrico CusinoCopyright:
Formati disponibili
Sbobine Funzione Renale -
Fisiologia
Fisiologia Umana
Università degli Studi di Urbino Carlo Bo (UNIURB)
36 pag.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
23/04/2020
FUNZIONE RENALE
Il rene è un organo tipicamente e prettamente omeostatico.
FUNZIONI DEL RENE (significati della funzione renale all’interno dell’organismo):
- Regolazione del pH e omeostasi degli ioni idrogeno: il rene ha una funzione importantissima nel
controllo e nel mantenimento del valore del pH. Se si blocca la funzione renale (blocco renale), si
muore principalmente per acidosi, infatti il pH raggiunge valori fuori controllo (inferiori a 7)
incompatibili con la vita.
- Omeostasi idrica: intesa sia come l’osmolarità totale del liquido extracellulare (rapporto totale tra
acqua e soluti) sia come il volume totale di acqua contenuto nel corpo.
- Eliminazione di cataboliti: per esempio l’urea è un catabolita (prodotto del catabolismo delle
molecole azotate), che come altri cataboliti vengono eliminati con l’urina.
- Omeostasi degli ioni: la concentrazione di ioni (a parte gli ioni idrogeno di cui se ne parla nel pH)
nel liquido extracellulare viene mantenuta ai valori corretti. La concentrazione di ioni come sodio,
potassio, calcio e fosfato deve essere regolata rigidamente per essere compatibile con la vita.
- Funzione endocrina: cioè la secrezione di ormoni (renina-angiotensina, eritropoietina, calcitriolo).
La funzione endocrina non è legata alla funzione omeostatica del rene (a parte la renina).
Le fasi attraverso cui si realizzano le prime 4 funzioni sopra elencate sono due. Le due fasi avvengono in
sequenza e in localizzazioni anatomiche diverse.
1. Filtrazione glomerulare.
La filtrazione consiste nella preparazione di una grande quantità di un liquido, chiamato
ultrafiltrato, molto simile al plasma.
2. Elaborazione del filtrato (nei tubuli).
La seconda fase consiste nell’elaborazione del liquido attraverso processi di riassorbimento
(sottrazione di soluti e di acqua) e di secrezione (aggiunta di soluti). La stragrande maggioranza di
acqua viene riassorbita, mentre alcuni soluti vengono assorbiti completamente e quindi non si
troveranno nel liquido finale (urina), altri soluti invece vengono riassorbiti solo in parte.
Il filtrato, o ultrafiltrato (prodotto della prima fase), somiglia molto al plasma, tranne per il fatto che non
presenta proteine. La composizione dell’ultrafiltrato infatti presenta per la maggior parte acqua, poco
potassio, molto sodio e cloro, bicarbonato, eccetera. Nel plasma invece, oltre a queste componenti, c’è
anche un 8% di proteine.
L’urina prodotta in un giorno è di circa 1,5 o 2 litri ed è composta prevalentemente da acqua. Ma questa
quantità da quanto filtrati deriva, cioè quanta acqua è stata riassorbita?
Ogni giorno si produce circa 180 litri di filtrato. Quel litro e mezzo di urina deriva da quei 180 litri, dunque
più del 99% di acqua del filtrato è stata riassorbita.
Nella colonna più a destra è indicata la percentuale di quanto è stato sottratto o aggiunto lungo il percorso
nei tubuli renali (nella seconda fase), durante il processo di elaborazione del filtrato.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
La percentuale di acqua restante è 0.6%.
Del sodio ne è rimasto lo 0.4-0.6%
Il potassio viene riassorbito in modo molto variabile in base a quanto ne introduciamo con
l’alimentazione.
Il calcio e il cloruro sono stati in massima parte riassorbiti.
Il bicarbonato ne rimane quasi niente (in condizioni normali).
Di glucosio, come amminoacidi e altre sostanze utili se ne trovano circa 0: tutto il glucosio viene
riassorbito, in quanto substrato energetico.
Le proteine erano presenti in quantità molto bassa e vengono comunque quasi interamente
riassorbite, ne rimane il 3%. Di urea ne resta circa metà.
L’ultimo valore è di 3000 in quanto, nell’urina non solo troviamo il 100% degli ioni idrogeno che
sono stati filtrati, ma ne troviamo 30 volte di più. Ciò indica che gli ioni idrogeno non solo vengono
filtrati nella prima fase della funzione renale ma vengono anche aggiunti in grande quantità.
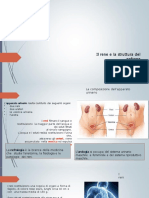
ANATOMIA FUNZIONALE
L’unità funzionale, cioè la più piccola parte del rene in grado di svolgere la funzione renale (produrre urina),
è il nefrone. Un altro aspetto anatomico fondamentale per comprendere la funzione del rene è la
circolazione renale.
I reni sono 2, si trovano nella parte cefalica e dorsale della cavità addominale, per cui sono appoggiati alla
parete posteriore della cavità addominale e sono subito sotto il diaframma.
I reni ricevono il sangue dall’arteria renale che è un ramo dell’aorta addominale, il sangue poi esce dalla
vena renale e si getta nella vena cava inferiore.
Il rene ha una forma a grosso fagiolo con una faccia convessa e una concava. La faccia concava si chiama ilo
e attraverso questo entrano l’arteria renale e i nervi ed escono l’arteria renale, i vasi linfatici e l’uretere.
L’uretere è un lungo tubo la cui parete ha una natura muscolare liscia, rivestito da un epitelio intero.
È molto lungo perché dall’ilo attraversa tutta la lunghezza della cavità addominale e si va a gettare nella
vescica.
La vescica è un organo cavo, la cui parete è prevalentemente di natura muscolare liscia, nella vescica
sboccano i due ureteri dx e sx e da essa caudalmente esce un altro tubo, l’uretra, verso l’esterno.
L’uretra è molto diversa nel maschio e nella femmina: nella femmina l’uretra sbocca nella vulva, tra piccole
e grandi labbra; nel maschio invece l’uretra è molto più lunga perché percorre tutta la lunghezza del pene e
inoltre confluisce con la via di espulsione dello sperma. Nell’uomo c’è un unico tubo terminale che sbocca
all’esterno che appartiene sia all’apparato genitale che a quello urinario.
Nella donna è più frequente la cistite (infezione della vescica) in quanto essendo più corta l’uretra, la
vescica è collegata all’esterno in modo più diretto rispetto all’uomo.
Nel rene si distinguono una parte interna o midollare, e una parte esterna o corticale.
La parte midollare è divisa in piramidi che hanno il vertice che guarda verso l’interno e che confluisce in
strutture tubulari molto corte che confluiscono in una cisterna, chiamata pelvi renale.
CIRCOLAZIONE RENALE
Il rene è un organo abbastanza piccolo e nonostante questo, l’arteria renale è molto grossa, infatti il 20%
del totale del sangue della gittata cardiaca va a finire nel rene.
L’arteria renale entra nell’ilo e si divide in rami che entrano nella midollare tra una
piramide renale e l’altra. I rami che passano tra le piramidi si chiamano arterie
interlobari, in quanto le piramidi renali si chiamano anche lobi.
Le arterie interlobari arrivate al confine tra midolllare e corticale, l’arteria interlobare
piega a 90° e percorre la superficie di base della piramide (quindi il confine tra midollare
e corticale). Siccome le basi delle piramidi sono curve, le arterie che le percorrono si
chiamano arciformi (a forma di arco). Dalle arterie arciformi partono delle arterie più
piccole, chiamate interlobulari, che salgono nella corticale dividendola in tanti settori
chiamati lobuli. Dalle arterie interlobulari si staccano delle arteriole afferenti, per cui c’è
un passaggio netto della componente istologica del vaso: nelle arterie c’è una prevalenza
di tessuto elastico, ora nelle arteriole c’è una prevalenza di tessuto muscolare liscio.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Le arteriole afferenti si dividono in capillari e portano sangue ad un gomitolo di capillari chiamato
glomerulo renale. Il glomerulo è una struttura vascolare fatta di capillari in rete che ricevono il sangue
dall’arteriola afferente.
A questo punto succede una cosa che succede solo in questo punto del corpo, tanto che è stata definita
rete mirabile: la rete di capillari, invece che riunirsi in una venula, si riunisce in una arteriola efferente.
Sia l’arteriola afferente che quella efferente hanno una grossa capacità contrattile.
L’arteriola efferente da origine ad una seconda rete capillare che va prevalentemente nella midollare
circondando altre strutture. I capillari si riuniscono poi in venule che sboccano in vene arciformi che si
riuniscono in vene interlobari, che si riuniscono nella vena renale.
Seguenza ricapitolativa: arteria renale arterie interlobari arteria arciformi arterie interlobulari
arteriole afferenti (+ grossa) capillari arteriole efferenti (+ piccola) capillari venule vene
arciformi vene interlobari vena renale.
Il glomerulo è circondato da una struttura chiamata capsula di Bowman. La capsula di Bowman è una
strutta doppia, una membrana (simile alla pleura) dentro cui si è infilato il glomerulo.
Per cui la capsula di Bowman presenta un foglietto interno (viscerale) che riveste direttamente il
glomerulo, e un foglietto esterno (parietale). Tra i due foglietti è presente lo spazio della capsula di
Bowman dentro cui si raccoglie il filtrato glomerulare.
Dentro il foglietto parietale della capsula di Bowman origina un tubo che ha un andamento molto sinuoso,
chiamato tubulo contorto prossimale facendo dei giri su se stesso; questo successivamente diventa una
tubo dritto che si dirige verso la midollare del rene (tubulo discendente).
I tubuli che originano più profondamente nella corticale, arrivano più profondamente nella midollare,
mentre invece i tubuli che originano più superficialmente nella corticale, arrivano più superficialmente nella
midollare.
Il tubo discendente drittissimo che punta verso la pelvi renale, va nella midollare arrivando a pronfondità
diverse, forma una U e torna su dritto, formando il tubo ascendente che decorre parallelo al tubo
discendente. La curva ad U che si forma si chiama ansa di Henle.
I due tubi sono chiamati rispettivamente braccio diascendente e braccio ascendente dell’ansa di Henle. Il
braccio ascendente presenta un primo tratto sottile con un epitelio piatto, dopodiché la parete si ispessisce
a causa dell’ispessimento dell’epitelio.
Arrivato nella corticale il tubulo riprende un andamento sinuoso, e viene chiamato tubulo contorto distale.
Il tubulo contorto distale, ad un certo punto del suo percorso va a passare esattamente a cavallo tra
arteriola afferente e arteriola efferente del corpuscolo renale da cui è nato il tubulo stesso.
Il corpuscolo renale è l’insieme del glomerulo e della capsula di Bowman.Tanti tubuli contorti distali infine si
gettano in un tubulo più grosso, molto dritto, chiamato dotto collettore (collettore, perché colleziona tanti
tubuli contorti distali, il dotto collettore non appartiene ad un solo nefrone ma è condiviso da tanti nefroni)
che sbocca nell’apice della piramide che poi riversa il contenuto nella pelvi renale.
I tubuli renali sono circondati da capillari peritubulari, che originano dall’arteria efferente. Essi raccolgono
ciò che esce dai tubuli renali (solventi e soluti); drenano lo spazio extracellulare. Dalla rete di capillari
peritubulari originano dei capillari estremamente lunghi che si dirigono in profondità nella midollare;
seguono il percorso delle anse di Henle. Hanno anche loro un andamento ad ansa, ed essendo una rete
parallela alla rete i capillari peritubulari, sono poco vascolarizzati. Inoltre, secondo Poiseuille, essendo
lunghi, hanno un alta resistenza al passaggio di sangue e il poco sangue che ci passa è molto lento. Questi
vasi capillari si chiamano vasa recta perché si dirigono dritti senza formare “pieghe” e seguono il percorso
dell’ansa si Henle. Anche questi sono importanti sempre per una funzione di drenaggio e di mantenimento
dell’ambiente extracellulare della midollare.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il
glomerulo presenta un’arteriola afferente (quella più grande), e una efferente
(quella più piccola). L’arteriola afferente si divide in tanti capillari che si
riuniscono nell’arteriola efferente. Il foglietto pariteale della capsula di
Bowman (tale capsula è lo slargamento del tubulo contorto prossimale) è
costituito da un epitelio paviementoso, e nel punto di contatto con le arteriole
(afferente ed efferente), si ripiega su se stesso e forma il foglietto viscerale,
che riveste ciascun signolo capillare glomerulare.
Il foglietto viscerale è fatto di podociti, cellule che hanno un corpo cellulare
(dove ci sono il nucleo e la maggior parte degli organuli) e dei prolungamenti
molto lunghi che danno origine a dei rami che sono a loro volta ramificati fino
a finire in piedini terminali, chiamati pedicelli.
I piedi sono le ramificazioni terminali dei rami del podocita. I rami del podocita
si intrecciano l’uno con l’altro, formando una struttura che ha tanti spazi
intercellulari tra un piedino e l’altro, ma senza spazi vuoti: i piedi infatti sono
intersecati fittamente l’uno con l’altro lasciando solo un piccolo spazio vuoto. Tutto il capillare è rivestito di
questo intreccio di piedi.
Nel capillare (in fig. a dx) sta passando un eritrocita, per
cui l’ingrandimento dell’immagine è molto forte. Le
cellule endoteliali del capillare sono legate alla lamina
basale (rete tridmensionale di proteine, è una struttira
extracellulare). Alla lamina basale sono attaccati i
pedicelli del podociti.
Tra i pedicelli sono visibili gli spazi che ci sono tra uno e
l’altro.
L’endotelio del capillare è di tipo fenestrato, cioè in certi
punto la membrana esterna della cellula endoteliale si
ripega su quella intena formando dei fori piuttosto
grandi, e pieni di proteine. Tra i fori dell’endotelio
capillare e tra gli spazi dei pedicelli possono passare
bene anche molecole piuttosto grandi.
La lamina basale invece è formata da una rete molto stretta e fitta, che difficilmente lascia passare le
molecole più grandi.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
24/04/2020
Il glomerulo, la capsula di Bowman e tutto l’insieme dei tubuli costituiscono l’unità funzionale del rene,
cioè il nefrone. Ovviamente il dotto collettore non appartiene ad un solo nefrone, ma è condiviso tra tanti
nefroni. Nel nefrone si distingue una parte vascolare (glomerulo) e una parte urinifera (insieme dei tubuli
dove si forma l’urina).
L’arteriola afferente (9), entra nella capsula di Bowman, si sfiocca nei capillari (10) che sono stati tagliati
trasversalmente, e che a loro volta si riuniscono nella via efferente (11) che porta via il sangue. Il tubulo
contorto distale (C) è stato anch’esso tagliato trasversalmente e passa tra arteriola afferente e arteriola
efferente. All’interno del glomerulo, ci sono i capillari con il loro endotelio, ma anche delle cellule che
stanno tutte intorno ai capillari, chiamate cellule del mesangio (“mesos-angios”= in mezzo ai capillari).
Le cellule del mesangio hanno delle capacità contrattili (non sono cellule muscolari). Esse hanno dunque la
capacità di ridurre il diametro del capillare, quindi di ridurre la superficie laterale del capillare.
Il tubulo contorto distale passa a cavallo tra l’arteriola afferente e quella efferente, in particolare passa
aderente alla parete dell’arteriola afferente (che ovviamente ha il suo endotelio e cellule muscolari lisce
come tutte le arteriole). Nel punto di contatto tra il tubulo contorto distale (che fa parte della parte
urinifera del nefrone) e l’arteriola afferente (che fa parte della parte ematica del nefrone), le cellule
epiteliali del tubulo contorto distali sono cellule specializzate, chiamate nel complesso macula densa (7).
Sono chiamate così in quanto al microscopio si vede una macchia di cellule più scure (perché assorbono di
più la luce). Fuori dal tubulo contorto distale, fuori dalla macula densa, si trovano delle altre cellule
specializzate (6), che stanno a ridosso della parete dell’arteriola, chiamate iuxta glomerulari (= vicino al
glomerulo). Nel complesso, poiché le cellule della macula densa e le cellule iuxta glomerulari, formano
un’unità funzionale, ci sono le caratteristiche per dire che insieme formano un apparato, cioè un insieme di
organi che formano un apparato che si chiama apparato iuxta glomerulare.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
FILTRAZIONE GLOMERULARE
È la prima fase della funzione renale.
La filtrazione è una separazione che avviene tra un materiale, un filtro e una forza che spinge il materiale
contro il filtro (forza di gravità).
Il materiale di partenza è il sangue all’interno dei capillari glomerulari (plasma contenente proteine, globuli
rossi, globuli bianchi e piastrine).
Il filtro è dato dalla barriera di separazione tra sangue e spazio della Capsula di Bowman (foglietto viscerale
e parietale) in altre parole dalla struttura di interfaccia dei capillari e dei pedicelli del foglietto viscerale,
questa barriera di separazione è costituita da tre strati: il primo e l’ultimo sono dei filtri a maglie larghe
(spazio tra pedicelli e capillari del glomerulo), mentre lo strato al centro, fa la differenza in quanto è un
filtro a maglie strette.
Attraverso il filtro, per cui passano tutte le strutture presenti nel sangue, eccetto che quelle più grandi, per
cui la parte corpuscolata: globuli rossi, globuli bianchi, piastrine. Inoltre vengono trattenute anche la
maggior parte delle proteine plasmatiche, vengono trattenute quelle più grandi di 65mila Dalton (è il limite
di dimensioni per il passaggio attraverso il filtro).
Le albumine, che sono le proteine più piccole tra quelle del plasma, hanno dimensioni tra 65 e 70 mila
dalton; ciò vuol dire che tutte le altre proteine che sono più grandi, verranno trattenute nel filtro. Le
albumine non passano tutte, ma solo quelle più piccole.
La lamina basale è fatta a sua volta di proteine che sono portatrici di carica elettrica (le proteine infatti
hanno gruppi che si ionizzano), quindi per determinare il passaggio o meno della proteina è importante
anche il tipo carica elettrica della proteina che passa. In piccola quantità inoltre ci sono proteine più piccole
delle albumine come per esempio l’insulina che ha un peso introno a 5 mila Dalton.
Le proteine che passano e arrivano nel filtrato glomerulare, in realtà non si ritrovano nell’urina, in quanto
vengono riassorbite prima di arrivare in fondo.
Quindi nello spazio della capsula di Bowman si va a formare un liquido privo, o quasi privo di proteine . Nel
filtrato della capsula sono presenti invece tutti gli altri componenti (ioni, glucosio, amminoacidi, ecc.).
Il liquido per passare attraverso la barriera filtrante incontra un attrito, per cui è necessario che ci sia una
forza che spinge.
Solitamente i meccanismi possibili di passaggio sono 2: quello di diffusione e quello di filtrazione. In questo
caso però quello di diffusone non ha importanza in quanto il materiale che passa ha circa la stessa
composizione del materiale di partenza: il filtrato glomerulare ha la stessa concentrazione della quasi
totalità delle strutture del plasma (l’unico componente diverso sono le proteine).
Il meccanismo che in questo caso entra in gioco è quello di filtrazione.
Nel meccanismo di filtrazione si devono considerare la pressione idrostatica (del sangue nel capillare
glomerulare; del filtrato nello spazio della capsula) e la pressione oncotica (del sangue nel capillare; del
filtrato nello spazio capsulare).
- La pressione oncotica del plasma (πc) è 25mmHg, poiché la concentrazione è sempre la stessa in
tutto il sangue. Questa pressione tenderà a trattenere l’acqua all’interno dei capillari.
- La pressione oncotica del filtrato nello spazio capsulare (πB) è praticamente 0mmHg, poiché di
proteine ne passano talmente poche che la pressione può essere approssimata a zero.
- La pressione idrostatica nei capillari tissutali è di 35mmHg e poi diminuisce fino a 15mmHg, per cui
possiamo considerare che la pressione media di un capillare (tolta la componente gravitazionale)
25mmHg.
Nei capillari glomerulari c’è una pressione idrostatica molto più grande rispetto a quella di
qualunque altro capillare di circa 48mmH.
- La pressione idrostatica del filtrato nella capsula di Bowman deve essere considerata sia in
relazione alla velocità di filtrazione, sia all’attrito del filtrato nei tubuli. La velocità di filtrazione
determina il flusso del liquido nella capsula e la spinta del flusso stesso all’interno dei tubuli. Per cui
se c’è un flusso importante e una resistenza alta, si ha un ΔP con un certo valore, diverso da zero.
La ΔP rappresenta differenza di pressione tra la capsula e la pelvi renale (nella pelvi renale c’è
pressione 0, poiché non c’è attrito). Il filtrato che si va a formare nella capsula di Bowman a seguito
della filtrazione prosegue lungo l’unica via di uscita, cioè lungo i tubuli, dove incontra un certo
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
attrito. Nella capsula di Bowman si forma una certa pressione che permette al liquido di essere
spinto attraverso i tubuli. Questa pressione ha un valore tra i 10 e i 15mmHg.
La pressione idrostatica nel capillare glomerulare spinge sulla parete del capillare e tende a far uscire il
liquido nello spazio della capsula di Bowman. La pressione idrostatica nella capsula di Bowman spinge il
liquido a rientrare nei capillari, per cui si oppone alla filtrazione del sangue. La pressione oncotica
all’interno del sangue nei capillari tende a trattenere acqua nei capillari, per cui anche questa si oppone alla
filtrazione.
La sommatoria tra le pressioni (48-12-25=11mmHg) danno una risultante di 11mmHg che rappresenta
l’effettiva pressione di filtrazione. È la pressione, la forza, che effettivamente permette la filtrazione.
Il valore della pressione non è molto alto; se diminuisse, il rene non funzionerebbe più, perché si avrebbe
l’interruzione della filtrazione renale, e quindi il blocco renale (=morte).
Un aumento della pressione oncotica (quindi una diminuzione della pressione totale) potrebbe avvenire
nella condizione in cui ci sono troppe proteine: ad esempio nel caso nel caso del doping si ha
un’iperfunzione del fegato che produce troppe proteine.
Nella condizione di cancro al fegato, cioè la produzione incontrollata di cellule epatiche, si ha una parte del
fegato in più (malata), che produce proteine plasmatiche determinando anche in questo caso
un’iperfunzione apatica.
In condizioni fisiologiche, la pressione oncotica resta costante (perché resta costante la concentrazione di
proteine plasmatiche); la pressione idrostatica della capsula di Bowman anche resta costante, perché è
determinata dalla lunghezza e dei tubuli, perciò da fattori anatomici; ciò che potrebbe variare è la pressione
idrostatica nei capillari glomerulari.
Se avessimo una caduta di pressione sistemica (per esempio dopo una seduta di esercizi, in cui si ha ancora
una vasodilatazione), si avrebbe anche una caduta di pressione glomerulare. Una diminuzione della
pressione arteriosa può essere permessa ovunque, ma non nel glomerulo perché se si perdono 10mmHg di
pressione glomerulare, si va in blocco renale.
Per evitare questa situazione ci sono dei meccanismi che permettono di controllare effettivamente la
pressione glomerulare (che è particolarmente alta rispetto agli altri capillari dell’organismo).
La pressione oncotica dei capillari glomerulari può assumere valori superiori ai 25mmHg, fino a 35mmHg.
Ciò avviene in quanto nel sangue aumenta la concentrazione di proteine scorrendo lungo il capillare, in
quanto le proteine vengono mantenute e l’acqua viene filtrata: a mano a mano che si procede lungo il
capillare la perdita d solvente fa si che la concentrazione di proteine aumenti. Quindi complessivamente si
ha un aumento della pressione oncotica dei capillari.
Se la pressione oncotica aumenta, la pressione di filtrazione totale diminuisce procedendo lungo il capillare.
In ascissa viene indicato la lunghezza del capillare; in ordinata si ha la
pressione effettiva di filtrazione.
La linea tratteggiata rappresenta la pressione idrostatica del sangue nel
capillare (rimane costante, diminuisce solo di un po’).
Anche la pressione della capsula di Bowman rimane costante all’incirca.
Ciò che cambia è la pressione oncotica che aumenta fino ad eguagliare la
pressione della capsula. Nel momento in cui la pressione oncotica
raggiunge quella della capsula di Bowman si azzera la pressione totale di
filtrazione. Per cui solo la prima metà del capillare glomerulare si impegna
nella formazione del filtrato.
Il risultato di tutto ciò è la produzione di liquido filtrato fino a 180 litri al giorno. Ovviamente non abbiamo
questa quantità di liquidi nel sangue e nel corspo; ciò sinifica che nel corpuscolo renale la parte liquida del
sangue (plasma) viene filtrato e riassorbito più volte al giorno.
Il volume del sangue totale è circa 5 litri, di questi circa 3 litri sono plasma. Questi 3 litri vengono filtrati
tante volte fino a raggiungere un volume totale di 180litri.
In realtà si devono considerare altri fattori, ma si può dire che se il plasma passasse interamente (3 litri alla
volta) attraverso il filtro, il plasma verrebbe filtrato e riassorbito 60 volte in un giorno. In realtà non è 60
volte, ma molto di più, perché viene filtrato solo una parte del plasma alla volta.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Chi ha un’insufficienza renale ed è costretto a fare dialisi tante volte alla settimana a seconda del grado di
insufficienza renale.
23/04/2020
FUNZIONE RENALE
Il rene è un organo tipicamente omeostatico.
FUNZIONI DEL RENE (significati della funzione renale nell’organismo):
- Eliminazione di cataboliti. L’urea è un catabolita, che come altri cataboliti vengono eliminati con
l’urina.
- Omeostasi idrica: intesa sia come l’osmolarità totale del liquido extracellulare (rapporto tra acqua
e soluti) sia come il volume totale di acqua contenuto nel corpo.
- Regolazione del pH (ioni idrogeno).
- Omeostasi degli ioni. La concentrazione di ioni (a parte gli ioni idrogeno di cui se ne parla nel pH)
nel liquido extracellulare viene mantenuta ai valori corretti. Ioni come sodio, potassio, fosfato. La
concentrazione deve essere regolata rigidamente per essere compatibile con la vita.
- Funzione endocrina, cioè la secrezione di ormoni (renina-angiotensina; eritropoietina; calcitriolo).
Se si blocca la funzione renale (blocco renale), si muore principalmente per acidosi: il pH raggiunge valori
fuori controllo (inferiori a 7) incompatibili con la vita.
Le fasi attraverso cui si realizzano le prime 4 funzioni sopra elencate sono due:
3. Filtrazione glomerulare
4. Elaborazione del filtrato (nei tubuli)
Le due fasi avvengono in sequenza e in localizzazioni anatomiche diverse.
La filtrazione consiste nella preparazione di una grande quantità di un liquido, chiamato ultrafiltrato, molto
simile al plasma. La seconda fase consiste nell’elaborazione del liquido attraverso processi di
riassorbimento (sottrazione di soluti e di solvente) e di secrezione (aggiunta di soluti). La stragrande
maggioranza di acqua viene riassorbita, mentre alcuni soluti vengono assorbiti completamente e quindi non
si troveranno nel liquido finale (urina), altri soluti invece vengono riassorbiti solo in parte.
Il filtrato, o ultrafiltrato (prodotto della prima fase), somiglia molto al plasma, tranne per un fattore: non
presenta proteine. La composizione dell’ultrafiltrato presenta per la maggior parte acqua, poco potassio,
molto sodio e cloro, bicarbonato eccetera. Nel plasma invece, oltre a queste componenti, c’è anche un 8%
di proteine.
L’urina prodotta in un giorno è di circa 1,5 o 2 litri. L’urina è composta prevalentemente di acqua.
Ogni giorno si produce circa 180 litri di filtrato. Quel litro e mezzo di urina deriva da quei 180 litri, dunque
più del 99% di acqua del filtrato è stata assorbita.
Nella colonna più a destra è indicata la percentuale
di quanto è stato sottratto o aggiunto lungo il
percorso nei tubuli renali (nella seconda fase).
La percentuale di acqua restante è 0.6%.
Del sodio ne è rimasto lo 0.4-0.6%
Il potassio viene riassorbito in modo molto variabile
in base a quanto ne introduciamo con
l’alimentazione.
Il calcio e il cloruro sono stati in massima parte
riassorbiti.
Il bicarbonato ce ne rimane quasi niente (in
condizioni normali).
Di glucosio, come altre amminoacidi se ne trovano 0: tutto il glucosio viene riassorbito, in quanto substrato
energetico. Le proteine erano presenti in quantità molto bassa e vengono comunque quasi interamente
riassorbite, ne rimane il 3%. Di urea ne resta circa metà.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
L’ultimo valore è di 3000 in quanto, nell’urina non solo troviamo il 100% degli ioni idrogeno che sono stati
filtrati, ma ne troviamo 30 volte di più. Ciò indica che gli ioni idrogeno non solo vengono filtrati, ma anche
aggiunti.
ANATOMIA FUNZIONALE
L’unità funzionale, cioè la più piccola parte del rene in grado di svolgere la funzione renale (produrre urina),
è il nefrone. Un altro aspetto anatomico fondamentale per comprendere la funzione del rene è la
circolazione renale.
I reni sono 2, si trovano nella parte cefalica e dorsale della cavità addominale, per cui sono appoggiati alla
parete posteriore della cavità addominale e sono subito sotto il diaframma.
I reni ricevono il sangue dall’arteria renale che è un ramo dell’aorta addominale; il sangue poi esce dalla
vena renale che si getta nella vena cava inferiore.
Il rene ha una forma a grosso fagiolo con una faccia convessa e una concava. La faccia concava si chiama ilo
e attraverso questo entrano l’arteria renale e i nervi ed escono l’arteria renale, i vasi linfatici e l’uretere.
L’uretere è un lungo tubo la cui parete ha una natura muscolare liscia, ovviamente rivestito da un epitelio
intero. È molto lungo perché dall’ilo attraversa tutta la cavità addominale e si va a gettare nella vescica. La
vescica è un organo cavo, la cui parete è prevalentemente di natura muscolare liscia. Nella vescica
sboccano i due ureteri dx e sx e da essa caudalmente esce un altro tubo, l’uretra, verso l’esterno. L’uretra è
molto diversa nel maschio e nella femmina: nella femmina l’uretra sbocca nella vulva, tra piccole e grandi
labbra; nel maschio invece l’uretra è molto più lunga perché percorre tutta la lunghezza del pene e inoltre
confluisce con la via di espulsione dello sperma. Nell’uomo c’è un unico tubo terminale che sbocca
all’esterno che appartiene sia all’apparato genitale che a quello urinario.
Nella donna è più frequente la cistite (infezione della vescica) in quanto essendo più corta l’uretra, la
vescica è collegata all’esterno in modo più diretto rispetto all’uomo.
Nel rene si distinguono una parte più interna, o midollare, e una parte esterna, corticale. La parte midollare
è divisa in piramidi renali che hanno il vertice che guarda in strutture tubulari molto corte che confluiscono
in una cisterna, chiamata pelvi renale.
Il rene è un organo abbastanza piccolo e nonostante questo, l’arteria renale è molto grossa, infatti il 20%
del totale del sangue della gittata cardiaca va a finire nel rene. L’arteria renale entra
nell’ilo e si divide in rami che entrano nella midollare tra una piramide renale e l’altro.
I rami che passano tra le piramidi si chiamano arterie interlobari, in quanto le piramidi
renali si chiamano anche lobi. Arrivate le arterie interlobari al confine tra midolllare e
corticale, l’arteria interlobare piega a 90° e percorre la superficie di base della piramide
(quindi il confine tra midollare e corticale della piramide). Siccome le basi delle piramidi
sono curve, le arterie che le percorrono si chiamano arciformi (a forma di arco). Dalle
arterie arciformi partono delle arterie più piccole, chiamate interlobulari, che salgono
nella corticale dividendola in tanti settori chiamati lobuli. Dalle arterie interlobulari si
staccano delle arteriole afferenti, per cui c’è un passaggio netto della componente
istologica del vaso: nelle arterie c’è una prevalenza di tessuto elastico, ora nelle arteriole
c’è una prevalenza di tessuto muscolare liscio. Sono arteriole afferenti che portano
sangue ad un gomitolo di capillari che si chiama glomerulo renale. Il glomerulo è una
struttura vascolare fatta di capillari in rete che ricevono il sangue dall’arteriola afferente.
A questo punto succede una cosa che succede solo in questo punto del corpo, tanto che è stata definita
rete mirabile: la rete di capillari, invece che riunirsi in una venula, si riunisce in una arteriola efferente.
Sia l’arteriola afferente che quella efferente hanno una grossa capacità contrattile.
L’arteriola efferente da origine ad una rete di capillari che vanno prevalentemente nella midollare che
circonda altre strutture. I capillari si riuniscono poi in venule che sboccano in vene arciformi che si
riuniscono in vene interlobari, che si riuniscono nella vena renale.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il glomerulo è circondato da una struttura chiamata capsula di Bowman. La capsula di Bowman è una
strutta doppia, una membrana (un po’ come la pleura) dentro cui si è infilato il glomerulo.
Per cui la capsula di Bowman presenta un foglietto interno (viscerale) che riveste
direttamente il glomerulo, e un foglietto esterno (parietale). Tra i due foglietti è presente lo
spazio della capsula di Bowman dentro cui si raccoglie il filtrato glomerulare.
Dentro il foglietto parietale della capsula di Bowman origina un tubo che ha un andamento
molto sinuoso, chiamato tubulo contorto prossimale facendo dei giri su se stesso; questo
successivamente diventa una tubo dritto che si dirige verso la midollare del rene. I tubuli che
originano più profondamente nella corticale, arrivano più profondamente nella midollare; i
tubuli che originano più superficialmente nella corticale, arrivano più superficialmente nella
midollare. Il tubo discendente drittissimo che punta verso la pelvi renale, va nella midollare
arrivando a pronfondità diverse, forma una U e torna su dritto, formando il tubo ascendente
che decorre parallelo al tubo discendente. La curva ad U che si forma si chiama ansa di
Henle. I due tubi sono chiamati rispettivamente braccio diascendente e braccio ascendente
dell’ansa di Henle. Il braccio ascendente presenta un primo tratto sottile con un epitelio
piatto, dopodiché la parete si ispessisce a causa dell’ispessimento dell’epitelio. Arrivato nella
corticale il tubulo riprende un andamento sinuoso, e viene chiamato tubulo contorto distale. Il
tubulo contorto distale, ad un certo punto del suo percorso va a passare esattamente a cavallo
tra arteriola afferente e arteriola efferente del corpuscolo renale da cui è nato il tubulo stesso
(fig. a dx.).
Il corpuscolo renale è l’insieme del glomerulo e della capsula di Bowman.Tanti tubuli contorti
distali infine si gettano in un tubulo più grosso, molto dritto, chiamato dotto collettore
(collettore, perché colleziona tanti tubuli contorti distali) che sbocca nell’apice della piramide
che poi riversa il contenuto nella pelvi renale.
I tubuli renali sono circondati da capillari peritubulari, che originano dall’arteria efferente. Essi
raccolgono ciò che esce dai tubuli renali (solventi e soluti); drenano lo spazio extracellulare.
Dalla rete di capillari peritubulari originano dei capillari estremamente lunghi che si dirigono in
profondità nella midollare; seguono il percorso delle anse di Henle. Hanno anche loro un
andamento ad ansa, ed essendo una rete parallela alla rete i capillari peritubulari, sono poco
vascolarizzati. Inoltre, secondo Poiseuille, essendo lunghi, hanno un alta resistenza al passaggio
di sangue e il poco sangue che ci passa è molto lento. Questi si chiamano vasa recta perché si
dirigono dritti senza formare “pieghe” e seguono il percorso dell’ansa si Henle. Anche questi
sono importanti sempre per una funzione di drenaggio e di mantenimento dell’ambiente
extracellulare della midollare.
Il glomerulo presenta un’arteriola afferente (quella più grande), e una
efferente (quella più piccola).
L’arteriola afferente si divide in tanti capillari che si riuniscono
nell’arteriola efferente. Il foglietto pariteale della capsula di Bowman è
costituito da un epitelio paviementoso, e nel punto di contatto con le
arteriole, si ripiega su se stesso e forma il foglietto viscerale, che riveste
ciascun signolo capillare glomerulare. Il foglietto viscerale è fatto di
podociti, cellule che hanno un corpo cellulare (dove ci sono il nucleo e la
maggior parte degli organuli) e dei prolungamenti molto
lunghi che danno origine a dei rami che sono a loro volta
ramificati fino a finire in piedini terminali, chiamati pedicelli. I piedi sono le ramificazioni
terminali dei rami del podocita. I rami del podocita si intrecciano l’uno con l’altro, formando
una struttura che ha tanti spazi intercellulari tra un piedino e l’altro, ma non ci sono spazi
vuoti: i piedi sono intersecati fittamente l’uno con l’altro lasciando solo un piccolo spazio
vuoto. Tutto il capillare è rivestito di questo intreccio di piedi. (vedi fig. a dx).
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Nel capillare (in fig. in basso) sta passando un eritrocita, per cui
l’ingrandimento dell’immagine è molto forte. Le cellule endoteliali
del capillare sono legate alla lamina basale (rete tridmensionale di
proteine, è una struttira extracellulare). Alla lamina basale sono
attaccati i pedicelli del podociti.
Tra i pedicelli sono visibili gli spazi che ci sono tra uno e l’altro.
L’endotelio del capillare è di tipo fenestrato, cioè in certi punto la
membrana esterna della cellula endoteliale si ripega su quella intena
formando dei fori piuttosto grandi, e pieni di proteine. Tra i fori
dell’endotelio capillare e tra gli spazi dei pedicelli possono passare
bene anche molecole piuttosto grandi.
La lamina basale invece è formata da una rete molto stretta e fitta,
che difficilmente lascia passare le molecole più grandi.
Il glomerulo, la capsula di Bowman e tutto l’insieme dei tubuli costituiscono l’unità funzionale del rene, cioè
il nefrone. Ovviamente il dotto collettore non appartiene ad un solo nefrone, ma è condiviso tra tanti
nefroni. Nel nefrone si distingue una parte vascolare (glomerulo) e una parte urinifera (insieme dei tubuli
dove si forma l’urina).
L’arteriola afferente (9), entra nella capsula di Bowman, si
sfiocca nei capillari (10) che sono stati tagliati
trasversalmente, e che a loro volta si riuniscono nella via
efferente (11) che porta via il sangue. Il tubulo contorto
distale (C) è stato anch’esso tagliato trasversalmente e
passa tra arteriola afferente e arteriola efferente.
All’interno del glomerulo, ci sono i capillari con il loro
endotelio, ma anche delle cellule che stanno tutte intorno ai
capillari, chiamate cellule del mesangio (“mesos-angios”= in
mezzo ai capillari). Le cellule del mesangio hanno delle
capacità contrattili (non sono cellule muscolari). Esse hanno
dunque la capacità di ridurre il diametro del capillare,
quindi di ridurre la superficie laterale del capillare.
Il tubulo contorto distale passa a cavallo tra l’arteriola afferente e quella efferente; in particolare passa
aderente alla parete dell’arteriola afferente (che ovviamente ha il suo endotelio e cellule muscolari lisce
come tutte le arteriole). Nel punto di contatto tra tubulo contorto distale (che fa parte della parte urinifera
del nefrone) e arteriola afferente (che fa parte della parte ematica del nefrone), le cellule epiteliali del
tubulo contorto distali sono cellule specializzate, chiamate nel complesso macula densa (7). Sono chiamate
così in quanto al microscopio si vede una macchia di cellule più scure (perché assorbono di più la luce).
Fuori dal tubulo contorto distale, fuori dalla macula densa, si trovano delle altre cellule specializzate (6), che
stanno a ridosso della parete dell’arteriola, chiamate iuxta glomerulari (=vicino al glomerulo). Nel
complesso, poiché le cellule della macula densa e le cellule iuxta glomerulari, formano un’unità funzionale,
ci sono le caratteristiche per dire che insieme formano un apparato, cioè un insieme di organi che formano
un apparato che si chiama apparato iuxta glomerulare.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
FILTRAZIONE GLOMERULARE
È la prima fase della funzione renale.
La filtrazione è una separazione che avviene tra un materiale, un filtro e una forza che spinge il materiale
contro il filtro.
Il materiale di partenza è il sangue all’interno dei capillari glomerulari (plasma contenente proteine, globuli
rossi, globuli bianchi e piastrine).
Il filtro è dato dalla struttura di interfaccia capillari e pedicelli del foglietto viscerale, costituita da tre strati: il
primo e l’ultimo sono dei filtri a maglie larghe (spazio tra pedicelli e capillari del glomerulo), mentre lo
strato al centro, fa la differenza in quanto è un filtro a maglie strette.
Attraverso il filtro, per cui passano tutte le strutture presenti nel sangue, eccetto che quelle più grandi, per
cui la parte corpuscolata: globuli rossi, globuli bianchi, piastrine. Inoltre vengono trattenuti anche la
maggior parte delle proteine plasmatiche; vengono trattenute quelle più grandi di 65mila dalton (è il limite
di dimensioni per il passaggio attraverso il filtro). Le albumine, che sono le proteine più piccole tra quelle
del plasma, hanno dimensioni tra 65 e 70 mila dalton; ciò vuol dire che tutte le altre proteine che sono più
grandi, verranno trattenute nel filtro. Le albumine non passano tutte, ma solo quelle più piccole. La lamina
basale è fatta a sua volta di proteine che sono portatrici di carica elettrica (le proteine infatti hanno gruppi
che si ionizzano), quindi per determinare il passaggio o meno della proteina è importante anche il tipo
carica elettrica della proteina che passa. In piccola quantità inoltre ci sono proteine più piccole delle
albumine (come l’insulina che ha un peso introno a 5 mila dalton).
Le proteine che passano e arrivano nel filtrato glomerulare, in realtà non si ritrovano nell’urina, in quanto
vengono riassorbite prima di arrivare in fondo.
Quindi nello spazio della capsula di Bowman si va a formare un liquido privo, o quasi privo di proteine. Nel
filtrato della capsula sono presenti invece tutti gli altri componenti (ioni, glucosio, amminoacidi eccetera…).
Il liquido per passare attraverso la barriera filtrante incontra un attrito, per cui è necessario che ci sia una
forza che spinge.
Solitamente i meccanismi possibili di passaggio sono 2: quello di diffusione e quello di filtrazione. In questo
caso però quello di diffusone non ha importanza in quanto il materiale che passa ha circa la stessa
composizione del materiale di partenza: il filtrato glomerulare ha la stessa concentrazione della quasi
totalità delle strutture del plasma (l’unico componente diverso sono le proteine).
Il meccanismo che in questo caso entra in gioco è quello di filtrazione.
Nel meccanismo di filtrazione si devono considerare la pressione idrostatica (del sangue nel capillare
glomerulare; del filtrato nello spazio della capsula) e la pressione oncotica (del sangue nel capillare; del
filtrato nello spazio capsulare).
- La pressione oncotica del plasma (πc) è 25mmHg, poiché la concentrazione è sempre la stessa in
tutto il sangue. Questa pressione tenderà a trattenere l’acqua all’interno dei capillari.
- La pressione oncotica del filtrato nello spazio capsulare (πB) è praticamente 0mmHg, poiché di
proteine ne passano talmente poche che la pressione può essere approssimata a zero.
- La pressione idrostatica nei capillari tissutali è di 35mmHg e poi diminuisce fino a 15mmHg, per cui
possiamo considerare che la pressione media di un capillare (tolta la componente gravitazionale)
25mmHg.
Nei capillari glomerulari c’è una pressione idrostatica molto più grande rispetto a quella di
qualunque altro capillare di circa 48mmH.
- La pressione idrostatica del filtrato nella capsula di Bowman deve essere considerata sia in
relazione alla velocità di filtrazione, sia all’attrito del filtrato nei tubuli. La velocità di filtrazione
determina il flusso del liquido nella capsula e la spinta del flusso stesso all’interno dei tubuli. Per cui
se c’è un flusso importante e una resistenza alta, si ha un ΔP con un certo valore, diverso da zero.
La ΔP rappresenta differenza di pressione tra la capsula e la pelvi renale (nella pelvi renale c’è
pressione 0, poiché non c’è attrito). Il filtrato che si va a formare nella capsula di Bowman a seguito
della filtrazione prosegue lungo l’unica via di uscita, cioè lungo i tubuli, dove incontra un certo
attrito. Nella capsula di Bowman si forma una certa pressione che permette al liquido di essere
spinto attraverso i tubuli. Questa pressione ha un valore tra i 10 e i 15mmHg.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
La pressione idrostatica nel capillare glomerulare spinge sulla parete del capillare e tende a far uscire il
liquido nello spazio della capsula di Bowman. La pressione idrostatica nella capsula di Bowman spinge il
liquido a rientrare nei capillari, per cui si oppone alla filtrazione del sangue. La pressione oncotica
all’interno del sangue nei capillari tende a trattenere acqua nei capillari, per cui anche questa si oppone alla
filtrazione.
La sommatoria tra le pressioni (48-12-25=11mmHg) danno una risultante di 11mmHg che rappresenta
l’effettiva pressione di filtrazione. È la pressione, la forza, che effettivamente permette la filtrazione.
Il valore della pressione non è molto alto; se diminuisse, il rene non funzionerebbe più, perché si avrebbe
l’interruzione della filtrazione renale, e quindi il blocco renale (=morte).
Un aumento della pressione oncotica (quindi una diminuzione della pressione totale) potrebbe avvenire
nella condizione in cui ci sono troppe proteine: ad esempio nel caso nel caso del doping si ha
un’iperfunzione del fegato che produce troppe proteine.
Nella condizione di cancro al fegato, cioè la produzione incontrollata di cellule epatiche, si ha una parte del
fegato in più (malata), che produce proteine plasmatiche determinando anche in questo caso
un’iperfunzione apatica.
In condizioni fisiologiche, la pressione oncotica resta costante (perché resta costante la concentrazione di
proteine plasmatiche); la pressione idrostatica della capsula di Bowman anche resta costante, perché è
determinata dalla lunghezza e dei tubuli, perciò da fattori anatomici; ciò che potrebbe variare è la pressione
idrostatica nei capillari glomerulari.
Se avessimo una caduta di pressione sistemica (per esempio dopo una seduta di esercizi, in cui si ha ancora
una vasodilatazione), si avrebbe anche una caduta di pressione glomerulare. Una diminuzione della
pressione arteriosa può essere permessa ovunque, ma non nel glomerulo perché se si perdono 10mmHg di
pressione glomerulare, si va in blocco renale.
Per evitare questa situazione ci sono dei meccanismi che permettono di controllare effettivamente la
pressione glomerulare (che è particolarmente alta rispetto agli altri capillari dell’organismo).
La pressione oncotica dei capillari glomerulari può assumere valori superiori ai 25mmHg, fino a 35mmHg.
Ciò avviene in quanto nel sangue aumenta la concentrazione di proteine scorrendo lungo il capillare, in
quanto le proteine vengono mantenute e l’acqua viene filtrata: a mano a mano che si procede lungo il
capillare la perdita d solvente fa si che la concentrazione di proteine aumenti. Quindi complessivamente si
ha un aumento della pressione oncotica dei capillari.
Se la pressione oncotica aumenta, la pressione di filtrazione totale diminuisce procedendo lungo il capillare.
In ascissa viene indicato la lunghezza del capillare; in ordinata si ha la
pressione effettiva di filtrazione.
La linea tratteggiata rappresenta la pressione idrostatica del sangue nel
capillare (rimane costante, diminuisce solo di un po’).
Anche la pressione della capsula di Bowman rimane costante all’incirca.
Ciò che cambia è la pressione oncotica che aumenta fino ad eguagliare la
pressione della capsula. Nel momento in cui la pressione oncotica raggiunge
quella della capsula di Bowman si azzera la pressione totale di filtrazione.
Per cui solo la prima metà del capillare glomerulare si impegna nella
formazione del filtrato.
Il risultato di tutto ciò è la produzione di liquido filtrato fino a 180 litri al giorno. Ovviamente non abbiamo
questa quantità di liquidi nel sangue e nel corspo; ciò sinifica che nel corpuscolo renale la parte liquida del
sangue (plasma) viene filtrato e riassorbito più volte al giorno.
Il volume del sangue totale è circa 5 litri, di questi circa 3 litri sono plasma. Questi 3 litri vengono filtrati
tante volte fino a raggiungere un volume totale di 180litri.
In realtà si devono considerare altri fattori, ma si può dire che se il plasma passasse interamente (3 litri alla
volta) attraverso il filtro, il plasma verrebbe filtrato e riassorbito 60 volte in un giorno. In realtà non è 60
volte, ma molto di più, perché viene filtrato solo una parte del plasma alla volta.
Chi ha un’insufficienza renale ed è costretto a fare dialisi tante volte alla settimana a seconda del grado di
insufficienza renale.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
27/04/2020
Il problema di mantenimento della velocità di filtrazione è molto importante, in quanto una piccola
variazione del valore della pressione idrostatica del filtrato può portare anche alla morte.
Poiché la pressione di filtrazione è un valore molto importante e vitale, esso è regolato da una pluralità di
meccanismi di controllo.
La velocità di filtrazione glomerulare è la quantità di filtrato che si forma nell’unità di tempo in tutti i
glomeruli renali. Essa dipende da una costante (K), data dalle caratteristiche della membrana filtrante, ed è
direttamente proporzionale alla pressione di filtrazione totale (PF) e al flusso ematico renale (RBF).
La filtrazione dipende dal mantenimento della pressione di filtrazione; ma se immaginiamo di mantenere la
pressione di filtrazione e di interrompere il passaggio di sangue attraverso i capillari glomerulari, è chiaro
che la filtrazione si fermerebbe poiché una volta persa l’acuqa dal glomerulo, la pressione oncotica
all’interno del glomerulo diventerebbe più grande, tendendo a tratterene l’acqua e portando a zero la
pressione di filtrazione totale.
Quindi abbiamo la necessità sia di mantenere una pressione di filtrazione, sia un
flusso ematico renale.
La filtrazione del glomerulo può essere schematizzata in questo modo:
Il flusso di sangue (RBF) attraverso l’arteriola afferente (R A) entra nel
glomerulo, dove si divide in due parti: una parte del flusso totale del
sangue prosegue ed esce attraverso l’arteriola efferente (R E); una
parte del volume di sangue invece esce attraverso la barriera filtrante
(Rc) e viene filtrato. La quantità di quest’ultima nell’unità di tempo è
la velocità di filtrazione glomerulare (GFR).
La velocità di filtrazione può essere regolata tramite la regolazione delle due grandezze da cui dipende
(pressione di filitrazione e flusso ematico). Queste due, a loro volta, possono essere modificate e regolate
cambiando la resistenza che offre ciascuno dei tre elementi (arteriola afferente, barriera filtrante e arteriola
efferente) al passaggio di sangue.
Se costringo le arteriole afferenti, secondo la legge di Poiseuille, la resistenza di quel
vaso al passaggio del sangue aumenta alla quarta potenza (di 16 volte). Se costringo
tutte le arteriole afferenti dei reni, il sanue trova un’alta resistenza ed entrerà meno
sangue nel rene: diminuisce il flusso ematico renale.
Un’altra conseguenza è che nel glomerulo la pressione tenderà a diminuire.
Nel grafico vediamo che più aumenta la resistenza dell’arteriola afferente, più
diminuisce il flusso ematico renale e diminuisce anche la filtrazione.
Se si stringe l’arteriola efferente, aumentano le resistenze complessive del rene al flusso ematico,
ma essendo la costrizione a valle del glomerulo, è anche vero che, aumentando la resisteza
dell’arteriola efferente, aumenta il ΔP tra il glomerulo e la circolazione venosa, quindi
aumenta la pressione all’interno del glomerulo. In sostanza si ha una dimuizione del
flusso ematico renale complessivo, e auamento della pressione idrostatica all’interno
del glomerulo. Complessivamente i due effetti si compensano ma prevale l’aumento di
aumento di pressione. Quindi di fatto la velocità di filtrazione aumenta.
Nel grafico si vede che se aumento la resistenza dell’arteriola efferente, il fusso ematico
renale diminuisce, ma poiché aumenta la pressione idrostatica glomerulare, si ha un
aumento complessivodella velocità di filtrazione.
Quindi e costringo l’arteriola afferente i ha una diminuzione del flusso renale e della velocità di filtrazione;
se costringo l’arteriola efferente ho una diminuzione del flusso renale, ma anche un aumento della velocità
di filtrazione data dall’aumento della pressione glomerulare.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il sangue che si trova nel glomerulo ad una certa pressione e che scorre con un certo flusso viene
filtrato. La filtrazione dipende dalla velocità di filtrazione, dal flusso e dalla costante (K) data dalla
superficie di filtrazione e dalle caratteristiche della barriera filtrante. La superficie di filtrazione è la
superficie di tutti i capillari glomerulari. Si può agire sulla velocità di filtrazione anche agendo sulla
superficie di filtrazione: i capillari glomerulari devono essere rimpiccioliti.
Nel glomerulo di sono delle cellule che circondano i capillari glomerulari che hanno capacità
contrattili, chiamate cellule del mesangio. Queste si possono contrarre così da diminuire la
superficie di filtrazione. In questo modo aumenta la riesistenza della barriera di filtrazione.
Poiché è aumentata la resistenza della barriera di filtrazione, il sangue arrivato nel glomerulo,
proseguirà nell’arteriola efferente, piuttosto che essere filtrato.
MECCANISMI DI CONTROLLO SUI TRE FATTORI
I meccanismi di controllo sono diversi proprio perché è di fondamentale importanza prevenire una caduta
di pressione della velocità di filtrazione a livello renale, nonostante la pressione arteriosa del corpo subisca
grandi variazioni.
Tra i numerosi sistemi di controllo ricordiamo solo i principali che garantiscono il mantenimento della
funzione renale e quindi della vita:
- Risposta miogenica: è un meccanismo molto semplice di risposta del muscolo liscio alla sua stessa
distensione. Quando il muscolo liscio viene disteso, si aprono canali del calcio a controllo
meccanico che stanno nella membrana delle cellule del muscolo liscio e il muscolo si contrae.
Quando si ha un aumento di pressione arteriosa generale, aumenta anche la pressione nelle
arteriole afferenti. Questo aumento di pressione determina una distensione della parete delle
arteriole afferenti, e quindi anche delle sue cellule muscolari lisce, che tramite il meccanismo di
risposta miogenica, si contraggono e diminuiscono il diametro dell’arteriola.
Il meccanismo limita a valle l’aumento di pressione, mantenendo costante la pressione
glomerulare, nonostante un aumento di pressione arteriosa.
- Feedback tubulo-glomerulare: nel punto di contatto tra tubulo contorto distale e arteriola
afferente, ci sono delle cellule specializzate che formano l’apparato iuxta glomerulare (composto
dalle cellule della macula densa e dalle cellule iuxta glomerulari).
Nel tubulo contorto prossimale c’è un intensissimo riassorbimento di sodio. La quantità di sodio
che viene riassorbito dipende da quanto velocemente scorre il liquido nel tubulo contorto
prossimale. Il riassorbimento di sodio avviene per la presenza di trasportatori, che hanno una certa
velocità di funzionamento: se il liquido scorre rapidamente, hanno meno tempo per riassorbire il
sodio; se il liquido scorre lentamente, hanno più tempo e ne assorbe di più. All’uscita del tubulo
contorto prossimale c’è più o meno sodio a seconda della velocità di filtrazione (cioè di scorrimento
del liquido). La concetrazione di sodio nei tratti successivi del tubulo è un indice diretto della
velocità di filtrazione.
In caso di un aumento della velocità di filtrazione, si ha un aumento del sodio (e del Cloruro che lo
accompagna) nel tubulo contorto distale. L’aumento di sodio e cloruro nel tubulo contorto distare,
determina un aumento di cloruro nelle cellule della macula densa: il cloruro entra nelle cellule della
macula densa. Aumentando la concentrazione di cloruro queste cellule si depolarizzano in quanto il
cloruro è carico negativamente.
Il cloruro in condizioni normali è più concentrato fuori dalle cellule, per cui tende ad entrare per
gradiente di concentrazione portando cariche negative all’interno della cellula. Se la concentrazione
di Cloruro intracellulare aumenta, l’entrata di Cloruro diminuisce sempre per via del gradiente di
concentrazione. Il cloruro che entra, spalma cariche negative nella faccia interna della membrana e
quindi contribuisce ad aumentare la differenza di potenziale tra inteno ed esterno. Per cui se
diminuisce l’entrata di cloruro, diminuisce anche la differenza di potenziale. La stessa cosa avviene
se aumento il potassio extracellulare (diminuisce l’uscta di potassio e la cellula si depolarizza), o se
aumento il sodio intracellulare. Se cambio il gradiente, cambia il flusso di quello ione e questo ha
un effetto sul potenziale di membrana.
La depolarizzazione delle cellule della macula densa determina l’apertura dei
canali del calcio voltaggio dipendenti: il calcio entra. L’entrata di calcio
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
determina il rilascio di adenosina, per via dell’esocitosi. L’adenosina può agire
su diversi recettori accoppiati a proteina G, ma i recettori di tipo 1 e quelli di
tipo 2 hanno azioni opposte (interpretano in modo opposto il messaggio
portato dall’adenosina). I recettori di tipo 2 attivano l’adenilato ciclasi
determinando vasodilatazione; i recettori di tipo 1 attivano la proteina G i che
determina un’inibizione dell’adenilato ciclasi, una diminuzione del cAMP e
un’inibizione della PKA, determinando vasocostrizione, in piarticolare ciò
avviene nell’arteriola afferente che ha questo tipo di recettori.
Una vasocostrizione dell’arteriola afferente determina una diminuzione del flusso renale e quindi
della velocità di filtrazione.
- Angiotensina 2: l’angiotensina 2 fa parte di un sistema ormonale costituito da due stostanze, che
sono la renina e l’angiotensina. La renina è prodotta dalle cellule dell’apparato iuxta glomerulare, e
viene secreta all’interno dell’arteriola afferente. La renina non è un ormone di per sé, ma fa parte di
questo sistema ormonale. Essa è un enzima proteolitico estremamente selettivo che agisce
specificamente su una proteina abbastanza grande, una globulina plasmatica, chiamata
angiotensinogeno. La renina stacca un segmento terminale dell’angiotensinogeno, chiamato
angiotensina 1, formato da 10 amminoacidi. L’angiotensina 1 quindi viene formata all’interno del
sangue nel momento in cui viene secreta renina dalle cellule iuxta glomerulari, che agisce
sull’angiotensinogeno e stacca l’angiotensina 1. L’angiotensina 1 si trova nel sangue e viene
traspostata dal flusso ematico in tutto il corpo e raggiunge i capillari polmonari. L’endotelio dei
capillari polmonari presenta una proteina di membrana che è un enzima proteolitico, chiamato ACE
(Angiotensin Conventing Enzyme). L’ACE presenta il sito enzimatico rivolto verso l’esterno della
cellula endoteliale, cioè rivolto verso il sangue; quando passa l’angiotensina 1, l’ACE la intercetta e
la converte in angiotensina 2, stacando altri 2 amminoacidi. L’angiotensina 2 è un potente
vasocostrittore (infatti “angio”= arteriola, “tensina”= tensione) che mette in contrazione la parete
delle arteriole, determinando un aumento di pressione perché agisce su tutte arteriole del corpo.
L’angiotensina 2 agisce su un recettore a 7 passi accoppiato a
proteina Gq, che attiva il meccanismo della fosfolipasi C. La
fosfolipasi C va a idrolizzare un fosfolipide di membrana, il
fosfatidilinositolo 4,5 difosfato, liberando nel citosol
inositolo1,4,5 trifosfato (IP3). Quest’ultimo si lega ad un
recettore (che è un canale del calcio) della membrana del
reticolo endoplasmatico, determinando l’apertura del canale.
Quindi esce il calcio dal reticolo per gradiente di
concentrazione che va a legarsi alla calmodulina (CAM) che a
sua volta si lega alla MLCK, attivandola. La MLKC, fosforila la
miosina, determinando la contrazione del muscolo liscio.
Quindi l’angiotensina 2 determina la vasocostrizione a livello arteriolare in tutto il corpo e quindi un
aumento della pressione arteriosa. Per quanto riguarada il sistema circolatorio renale, l’arteriola
che risponde prevalentemente è l’arteriola efferente, poiché ha più recettori dell’angiotensina 2.
Quindi l’angiotensina 2 una volta convertita e attivata va a detemrinare una vasocostrizione
dell’arteriola efferente determinando un aumento della filtrazione.
Il rilascio della renina da parte dell’apparato
iuxtaglomerulare è dovuto alla presenza di due
condizioni: l’aumento della pressione arteriosa e la
concentrazione di sodio nel tubulo contorto distale.
Quando diminuisce la velocità di filtrazione, una
quantità maggiore di sodio viene riassorbita nel
tubulo contorto prossimale e ne resta meno nei
segmenti successivi del nefrone. La bassa
concentrazione di sodio nel tubulo contorto distale,
che è indice di una diminuita velocità di filtrazione, determina un aumento della secrezione di
renina, che determina a sua volta una costrizione dell’arteriola efferente che aumenta la filtrazione.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
- Ortosimpatico: un’attivazione ortosimpatica determina una diminuzione della velocità di
filtrazione. Le innervazioni ortosimpatiche rilasciano noradrenalina nelle pareti delle arteriole che
determina costrizione. Entrambe le arteriole (afferente ed efferente) rispondono all’ortosimpatico:
diminuisce il diametro di entrambe le arteriole, quindi diminuscisce il
flusso renale, perché aumenta la resistenza del rene al flusso ematico.
Non si ha invece una grande variazione della pressione glomerulare,
poiché entrambe le arteriole sono costrette.
L’attivazione ortosimpatica (in condizioni di stress, tipo allenamento)
determina una diminuzione della velocità di filtrazione. Non si ha necessariamente una diminuzione
della quantità di urina prodotta, perché questo dipende dai processi di riassorbimento dell’acqua.
- Volemia: è il volume totale del sangue.
Anche questo deve essere regolato, perché è uno dei fattori che determinano la pressione
idrostatica del sangue, sia arteriosa che venosa. Il circolo sanguigno complessivamente è un
contenitore chiuso, con pareti elastiche; più sangue c’è, più aumenta la pressione. Parlando della
circolazione venosa abbiamo visto che un aumento di volume del sangue non si distribuisce
uniformemente nelle varie parti del sistema cardiovascolare, ma preferibilmente la maggior parte si
va a collocare nella parte venosa perché le vene hanno una maggiore compliance, quindi sono
maggiormente dilatabili e possono incamenrare una quantità maggiore di sangue. Se il sangue
viene accumulato nelle vene, anche l’ultima parte della circolazione venosa (l’atrio destro) viene
dilatata. Un aumento di volemia infatti va letto nell’atrio destro. Questo accumulo di sangue
nell’atrio destro da origine al riflesso di Bainbridge (avviene grazie alla presenza di tensiocettori).
In questo caso nella parete dell’atrio destro sono
presenti delle cellule secernenti, che sono cellule
endocrine, e secernono un ormone: il peptide
natriuretico atriale (ANP). Questo ormone viene
secreto quando c’è un aumto di volemia, quindi
quando la parete di atrio destro viene messa in
tensione da un aumento di sangue (probabilmente il meccanismo di secrezione avviene per la
presenza di canali del calcio a controllo meccanico. Quindi quando queste cellule vengono distese
entra il calcio che va ad indurre la secrezione dell’ormone da parte delle vescicole).
L’ormone ANP ha diverse azioni: “natriuretico” indica che ha un’azione di aumento
dell’eliminazione urnaria di sodio; l’ormone natriuretico atriale ha anche la funzione di
vasodilatazione generale e di entrambe le arteriole (afferente ed efferente). La dilatazione di
entrambe le arteriole porta ad un aumento del flusso ematico renale poiché diminuiscono le
resistenze del rene al flusso di sangue. Aumentando il flusso renale aumenta anche la velocità di
filtrazione. Quindi viene filtrata una quantità maggiore di acqua e soluti, creando il presupposto per
eliminarli con l’urina. Ma non è comunque detto che poi venga eliminata una quantità maggiore di
acqua e soluti, perché questo dipenderà da quanta ne viene riassorbita. Essendoci inizialmente un
maggiore di acqua.
ELABORAZIONE DEL FILTRATO
Il filtrato che è stato prodotto in quantità enormi (fino a 180 litri al giorno) nella capsula di Bowman, viene
inviato nei tubuli, dove la composizione e la quantità del liquido vengono cambiate completamente.
Più del 99% del filtrato viene riassorbito completamente, il resto, l’urina, viene espulso. Alcuni soluti
presenti nel filtrato glomerulare non si troveranno più nell’urina, altri se ne troveranno in concentrazione
minore, ed altri ancora in concentrazione maggiore. Le quantità totali di sostanze che si ritrovano nell’urina,
rispetto alle quantità presenti nel filtrato, sono completamente diversi. La cratinina è un catabolita della
creatina, essa non viene quasi riassorbita, quindi la quantità totale è circa la stessa che si ritrova nel filtrato;
tutto il resto viene riassorbito in massima parte o totalmente. Gli ioni idrogeno invece li troviamo in
quantità molto maggiore nell’urina, rispetto a quelli che si trovano nel filtrato. Gli ioni idrogeno vengono
aggiunti lungo il percorso dei tubuli.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il tubulo contorto prossimale e dalla capsula di Bowman da cui parte, si trovano nella corticale del rene;
nella midollare invece continua il braccio discendente, l’ansa di Henle e il braccio ascendente, che arrivato
di nuovo alla corticale forma il tubulo contorto distale. Dei bracci dell’ansa di Henle presentano un tratto
spesso e uno sottile. Il tubulo contorto distale infine sbocca nel dotto collettore che ritorna nella midollare e
sbocca nella pelvi renale. La distinzione tra midollare e corticale non è solo morfologica, ma funzionale:
l’ambiente extracellulare della corticale è un normale ambiente extracellulare che ha i componenti del
liquido extracellulare, sia in quantità che in qualità, ha la stessa osmolarità del sangue. Nella midollare
invece c’è una osmolarità molto più grande (iperosmolarità) che aumenta all’aumentare della profondità; la
composizione dei soluti è la stessa qualitativamente, ma non quantitativamente.
Nel tubulo contorto prossimale si ha il riassorbimento di sodio molto intenso dato dalla pompa sodio
potassio. È un riassorbimento fondamentale, perché da questo derivano tutti gli altri, nel senso che il
trasporto attivo di sodio (grazie al meccaismo di trasporto primario pompa sodio-potassio) mette in moto
tutti gli altri riassorbimenti, prima di tutto quello dell’acqua per osmosi. Per mantenere l’equilibrio
elettrostatico, all’assorbimento del sodio (carica positiva), viene assorbito anche il cloruro (carica negativa).
C’è infine un riassorbimento di molecole per trasporto
attivo secondario: il gradiente di sodio creato dalla
pompa sodio potassio determina l’attivazione di
meccanismi di trasporto attivo secondario, sia nel senso
di riassorbimento (glucosio, amminoacidi, bicarbonato),
sia nel senso della secrezione (ioni idrogeno).
Nell’ansa di Henle si deve distinguere il braccio
discendente (che è sottile) e il tratto sottile del braccio
ascendente. In queste zone non avviene nulla, o quasi,
eccetto che il riassorbimento di acqua. Il riassorbimento
di acqua è sempre passivo, sempre per gradiente
osmotico. In natura non esiste un trasportatore di
acqua: l’acqua passa sempre per gradiente osmotico tramite i fori che trova. In questi tratti c’è un buona
permeabilità all’acqua e questa viene riassorbita, oltre a qualche ione sodio e cloruro. Completamente
cambia la situazione nel tratto spesso del braccio ascendente dell’ansa: esso diventa assolutamente
impermeabile all’acqua, ma c’è un potentissimo riassorbimento di sodio e cloruro. La pompa sodio-potassio
lavora moltissimo consumando molta ATP, determinando il riassorbimento di sodio, cloruro e potassio
Il tubulo contorto distale e il dotto collettore presentano una permeabilità dell’acqua variabile. La
variaibilità è controllata dall’ormone antidiuretico. Nel tubulo contorto distale riprendono gli assorbimenti
(sodio, cloruro, bicarbonato, calcio, fosfato) e le secrezioni (ioni idrogeno). Nel dotto collettore continuano i
riassorbimenti di sodio e cloruro, compare la possibilità di riassorbire urea e ancora una volta secreazione o
riassorbimento di ioni idrogeno. Nel tubulo contorto distale e nel dotto collettore avvengono la maggior
parte delle regolazioni fini: essi controllano precisamente che la quantità di riassorbimento dei soluti sia
quella giusta. Lo scopo non è quello di fare un’urina giusta, ma quello di mantenere le varie omeostasi, le
giute concentrazione del liquido extracellulare e dell’ambiente interno.
Nella figura sotto si mette in relazione la struttura dei vari tratti del tubulo con la funzione: guardando
l’epitelio si può capire qualcosa sulla sua funzione.
Il grafico rappresenta un’analisi energetica dei vari tratti del nefrone: è descritto il consumo energetico
dell’epitelio dei vari tratti del tubulo.
La prima immagine in alto a destra rappresenta una cellula epiteliale del tubulo contorto prossimale. È una
cellula cubica con microvilli molto sviluppati dalla parte verso il lume del tubulo. All’interno, il volume della
cellula cubica è occupato dai mitocondri (quei corpi neri). Queste caratteristiche citologiche vengono
mantenute lungo tutto il tubulo contorto prossimale, anche se il numero di mitocondri diminuisce. Nel
gradico vediamo che nel tubulo contorto prossimale (S1) il consumo di ATP è molto alto. La maggior parte
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
dell’ATP di queste cellule viene quasi per intero consumato dalla pompa sodio-potassio.
I mitocondri sono indispensabili in questo tratto in quanto la pompa sodio
potassio consuma molto ATP, e questo viene prodotto dai mitocondri.
I villi hanno la funzione di aumentare la superficie di riassorbimento e di
secrezione e di catturatre il maggior numero di molecole che attraversano
velocemente il tubulo contorto prossimale, per riassorbirle.
Al tubulo contorto prossimale segue la parte sottile dell’ansa di Henle
(braccio discendente e tratto sottile del braccio ascendente). L’epitelio è
appiattito, quasi pavimentoso, e presenta cellule povere di mitocondri; i villi
sono scomparsi. Tutto ciò significa che il metabolismo energetico è scarso,
con una scarsa attività della pompa-sodio potassio, quindi il
consume di ATP da parte della pompa è molto basso (nel
grafico indicato con ATL). Essendo scomparso l’orletto
spazzola (villi), gli scambi con il filtrato sono pochi e
prevalentemente passivi.
Nel tratto discendente, l’attività energetica è quasi nulla, nel
tratto ascendente l’epitelio è poco più presso e c’è la
presenza di qualche mitocondio in più.
Nella parte spessa del braccio ascendente dell’ansa di Henle
le cellule diventano cilindriche, con enormi mitocondri che riempiono tutto lo spazio della cellula. Ciò
significa che il metabolismo è molto attivo e la pompa sodio-potassio va al massimo. Non c’è la presenza
dell’orletto spazzola poiché il flusso del liquido è molto più lento rispetto a quello del tubulo contorto
prossimale, in quanto i 2/3 del volume totale sono stati riassorbiti nel tubulo contorto prossimale, e un
altro 15% del volume totale è stato riassorbito nell’ansa di Henle; essendo stato riassorbito l’85% circa del
liquido totale, il liquido scorre più lentamente e non si ha più la necessità di creare villi. Si ha comunque una
grande efficienza di riassorbimento.
Il tubulo contorto distale presenta cellule cubiche, con parecchi mitocondri. Sia nella parte spessa dell’ansa
di Henle, sia nel tubulo contorto distale, sia nel dotto collettore, ci sono giungioni intercellulari (tra cellula e
cellula epiteliale) molto strette che non lasciano spazio nemmeno ad una stupida piccola molecola di acqua.
Nel grafico è rappresentato con DCT.
Il dotto collettore presenta cellule cubiche di tre tipi differenti dal punto di vista funzionale.
28/04/2020
Tutte le modifiche che avvengono sia in termini quantitativi che qualitativi nel filtrato glomerulare,
avvengono per via della pompa sodio-potassio.
Il riassorbimento di acqua che avviene nel tubulo contorto distale e nel tubulo collettore viene chiamato
facoltatitvo, rispetto al grande riassorbimento di acqua che avviene nel tubulo contorto prossimale, e che
vinene detto obbligatorio.
L’assorbimento obbligatorio di acqua è quello che avvine a prescindere dal bisogno di acqua dell’organismo
(sia che sia disiratato, superidratato o normoidratato); avviene indipendentemente dallo stato di
idratazione. Gli assorbimenti facoltativi nel tubulo contorto distlale e nel tubul collettore avvengono più o
meno a seconda del bisogno dell’organismo di trattenere o di eliminare acqua.
[Il riassorbimento di acqua varia molto a seconda dell’età: in base all’età si ha un cambiamento continuo di
acqua presente all’interno del corpo. L’invecchiamento può essere visto come l’incapacità dei tessuti di
trattenere acqua.]
RIASSORBIMENTO DI SODIO
ll riassorbimento di sodio avviene sia nel tubulo contorto prossimale, che in altre zone del tubulo. Esso
avviene sostanzialmente con un meccanismo fondamentalmente uguale in tutte le parti.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Epitelio polarizzato= è un epitelio assorbente, dove c’è un trasferimento di sostanza da una
parte all’altra. Gli epiteli sono polarizzati nel senso che la membrana apicale, che guarda
verso il lume del tubulo (in figura la membrana ondulata) è diversa dalla membrana
basolaterale, quella che guarda verso la base e anche verso la membrana laterale (in figura
sono le linee continue laterale e sotto). Queste due membrane sono diverse nella loro
composizione proteica, quindi le proteine di membrana che si trovano nella membrana
apicale sono diverse da quelle che si trovano nella membrana basolaterale.
Com’è possibile che la membrana venga costruita diversamente da un lato e dall’altro lato
della cellula?
La membrana della cellula, come tutto nella vita, deve essere continuamente rinnovato, deve essere
sostituita.
Come si formano le proteine (sintesi proteica): dal DNA e dalla sequenza di nuocleotidi viene costruita la molecola di RNA
messaggero, che a seguito di alcune elaborazioni, viene trasferito nel citosol. Nel citosol i ribosomi si attaccano al RNA messaggero
che tripletta per tripletta (3 nucleotidi alla volta) aggiungono un amminoacido alla proteina che si sta formando. Questo vale sia
per le proteine di membrana, sia per le proteine destinate all’esocitosi, sia per le proteine citosoliche.
Come fa una proteina a finire nella membrana e l’altra a finitre nel citosol?
Le proteine di membrna sicuramente devono avere almeno una sequenza di amminoacidi idrofoba per stare nel doppio strato
fosfolipidico.
Ci sono due localizzazione dei robosomi: i ribosomi liberi e i ribosomi legati al reticolo endoplasmatico rugoso. I ribosomi liberi nel
citosol formano la proteina che a mano a mano che si forma e si allunga, si stacca dal ribosoma e va nel citosol. Le proteine invece
sintetizzate dal ribosoma legate al reticolo endoplasmatico rugoso, a mano a mano che si formano e che si forma la catena,
vengono inniettate all’interno del reticolo stesso. La proteina del ribosoma legato al reticolo, va a finire all’interno del lume del
reticolo. Se però una proteina sintetizzata dal ribosoma legato al reticolo, presenta una parte idrofoba, nel momento in cui viene
inniettata nella membrana del reticolo e passa la parte idrofoba, quella rimane incastrata all’interno della membrana del reticolo.
La proteina tutta idrofila, passa all’interno del lume del reticolo; quella che invece ha un pezzo della sequenza di amminoacidi
idrofoba, resta incastrata nella membrana del reticolo.
Nel reticolo queste proteine vengono elaborate, idrolizzate (eccetera…), dopodiché dal reticolo si formano per gemmazione delle
vescicole, delle esteroflessioni della membrana del reticolo. Le vescicole quindi che si vanno a formare sono rivestite della
membrana del reticolo, cioè quella membrana che si porta dietro quelle proteine che sono state sintetizzate dai ribosomi legato al
reticolo endoplasmatico rugoso. Questa vescicola va a fondersi con l’apparato di Golgi, dove le proteine vengono elaborte.
Dall’apparato di Golgi nascono per gemmazione, delle vescicole che si vanno a fondere per esocitosi con la membrana plasmatica.
A questo punto le proteine di quel pezzo di membrana vanno ad essere proteine di membrana plasmatica (provenienti dunque dai
ribosomi legato al reticolo). Le proteine invece contenute all’intenro della vescicola che va in esocitosi venogo secrete.
Continuamente nelle nostre cellule c’è un’aggiunta e un’eliminazione di membrana: le memrbane vengono
rinnovete poiché la vita è dinamica.Epitelio polarizzato significa che la membrana apicale è diversa da
quella basolaterale nella composizione proteica.
Ma come è possibile che ci siano proteine diverse da una parte e dall’altra?
Le vescicole che nascono dall’apparato di Golgi, non vanno a fondersi in un punto qualunque della
membrana, ma vengono indirizzate da un sistema molto complesso, che comprende dei binari di
citoscheletro, al punto preciso in cui devono esere fuse. C’è un indirizzamento delle vescicole a seconda
delle proteine di membrana della vescicola stessa. A seconda delle proteine di membrana delle vescicole,
queste verranno idirizzate e fondersi o con la membrana basolaterale, o con la membrana apicale. Questo
spiega l’origine della diversa composizione in proteine delle due membrane, ma sappiamo che le
membrane sono dei mosaici fluidi: i fosfolipidi si muovono e le proteine stesse vengono trascinate con loro
a meno che non ci siano dei vincoli (legami con il citoscheletro o con la matrice extracellulare) che limitano
il movimento delle proteine. Quindi se le proteine si muovessero liberamente, anche se ci sono due
membrane diverse, dopo un po’ dovrebbe esserci un rimescolamento. Ciò che impedisce il movimento
delle proteine in una cellula epiteliale polarizzata, mantenendo la diversa identità delle due membrane,
sono le tight junction. Nel punto di contatto tra due cellule adiacenti, ci sono delle giunzioni serrate (thight
junction) che sono delle collane di proteine di membrana che si agganciano con le proteine analoghe
dell’altra membrana. Le due membrane laterali di due cellule adiacenti sono tenute molto vicine. Questo
crea una specie di barriera tutt’intorno alla cellula che impedisce la migrazione delle altre proteine di
membrana al di là della bariera stessa. Essendo queste giunzioni presenti tutt’intorno alla cellula, ciò
impedisce che le proteine della membrana apcale vadano a finire nella membrana basolaterale e viceversa.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il riassorbimento del sodio avviene perché nella membrana basolaterale sono presenti tante copie della
pompa sodio-potassio. La pompa sodio-potassio porta all’esterno 3 ioni sodio e all’intenro 2 ioni potassio
per ogni molecola di ATP idrolizzata. Quindi prende il sodio intracellulare e lo trasporta nel liquido
extracellulare circostante, dove entrarà nel capillari sanguigni e verrà portato via. La pompa sodio potassio
impoverisce la cellula di sodio. Il potassio entra forzatamente nella cellula; ma in realtà nella membrana
basolaterlae ci sono dei canali del potassio sempre aperti, per cui in parte il potassio torna indietro dalla
stessa membrana e non viene accumulato all’interno della cellula.
La concentrazione di sodio intracellulare è estremamente bassa. Questo determina che rispetto al filtrato
che sta scorrendo nel tubulo contorto prossimale, la concentrazione di sodio intracellulare è molto più
bassa perché nel filtrato (che deriva dal plasma) c’è la stessa concentrazione di sodio del plasma, che è
quella extracellulare.
Nella membrana apicale si stabilisce un gradiente di concentrazione e di potenziale di sodio che tende a far
entrare il sodio nella cellula. Nel momento in cui il sodio (che era contenuto nel filtrato) entra nella cellula
attraverso la membrana apicale, viene portato fuori dalla cellula tramite la membrana basolaterale.
Come fa ad entrare il sodio del filtrato all’interno della cellula?
Gli ioni non possono passare attraverso il doppio strato fosfolipidico, ma devono
avere o dei canali o dei trasportatori.
Il sodio presenta diverse vie attraverso cui entrare nella cellula:
1. Nella membrana apicale ci sono dei canali del sodio sempre aperti che
permettono l’entrata di sodio.
2. Ci sono dei meccanismi di cotrasporto, cioè dei meccaismi che
accoppiano l’entrata del sodio secondo gradiente, con l’entrata o l’uscita
di qualcosaltro (meccanismi di sinporto o di antiporto). In particolare nel
tubulo contorto prossimale c’è un meccanismo di simporto sodio-
cloruro, che utilizzano il gradiente di sodio, fa entrare forzatamente il
cloruro all’interno della cellula.
3. È presente poi un altro cotrasportatore: sodio-glucosio, che utlizzando il
grandiete di sodio, fa entrare anche il glucosio. Questo è presente in
gradi quantità tanto che nel breve tratto del tubulo contorto prossimale
tutto il glucosio viene portato via dal liquido tubulare.
4. Ci sono poi, sempre nelle membrana apicale, dei trasportatori sodio-amminoacidi. Poiché esistono
numerosi amminoacidi, anche i costrasportatori saranno differenziati per ciascuno gruppo di
amminoacidi (cotrasportatore sodio-amminoacidi bicarbossilici; cotrasportatore sodio-amminoacidi
biamminici e altri ancora per altri gruppi di amminoacidi).
Nella membrana apicale poi c’è un importante meccanismo di antiporto: antiporto sodio-ione idrogeno. E’
un meccanismo di antiporto: fa entrare il sodio secondo gradiente di potenziale elettrochimico che
determina l’ucita forzata di ioni idrogeno.
Tutte le particelle che entrano nella cellula, aumentano la loro concentrazione nella cellula e poi passano
attraverso la membrana basolaterale secondo un gradiente (quindi trasporto passivo), ognuno attraverso la
sua via. Il cloruro ad esempio ha dei canali sulla membrana basolaterale che gli permettono di uscire. Il
glucosio trova un trasportatore che permette il passaggio secondo gradiente (è una diffusione facilitata); è
una diffusione mediata da un trasportatore proteico di membrana che avviene secondo gradiente di
concentrazione. uscendo dalla cellula le molecole vengono poi catturate dal torrente ematico.
Tutti gli assorbimenti e secrezioni dipendono dal gradiente di sodio che si è formato a cavallo della
membrana apicale grazie all’azione della pompa sodio potassio che si trova esclusivamente solo nella
membrana basolaterale. Questo determina nel tubulo contorto prossimale diversi cambiamenti nella
concentrazione di diverse sostanze presenti nel filtrato rappresentiati nel grafico sotto.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
In ascissa è rappresentata la lunghezza del tubulo contorto prossimale, in ordinata la percentuale di quella
specifica sostanza a diverse distanze dall’inizio del tubulo contorto prossimale (X=0= capsula di Bowman).
Si è detto che nel tubulo contorto prossimale c’è un grandissimo riassorbimento
di sodio, per cui mi aspetterei di vedere nel grafico una diminuzione della
concentrazione di sodio nel liquido; questa resta però perfettamente costante (o
con una minima diminuzione). Se la concentrazione resta costante vuol dire che
per ogni sodio che è stato riassorbito, è stata riassorbita una quantità
proporzionale di acqua (solvente). Sodio e acqua nel tubulo contorto prossimale
vanno di pari passo.
Altri soluti diminuiscono, ciò vuol dire che vengono riassorbiti tramite i
meccanismi sopra descritti (ad esempio il fosfato, o il bicarbonato).
Ci sono altri soluti che scompaiono completamente nel corso del tubulo contorto
prossimale (glucosio, lattato, amminoacidi).
Il cloruro invece aumenta un po’ la sua concentrazione nel tubulo. Ciò vuol dire
che il cloruro non viene riassorbito nella stessa quantità dell’acqua riassorbita; poiché è stata riassorbita
una grande quantità di acqua, il cloruro di fatto aumenta la sua concentrazione. in quest’ultimo caso è stato
riassorbito più solvente che soluto.
Alla fine l’acqua riassorbita è circa i 2/3, per cui alla fine del tubulo contorto prossimane sono stati
riassorbiti circa 120 litri di 180 totali circa nell’arco delle 24h.
Di sodio ne viene riassorbito tanto perché ne viene filtrato tanto e perché ce n’è tanto
nel plasma.
Il sodio lo si assume con la dieta, poiché mangiando cibi, mangiamo anche il liquido
extracellulare, ricco di sodio. inoltre assumiamo sodio anche tramite il sale da cucina
(cloruro di sodio). ogni giorno assumiamo circa 120millimoli di sodio. Nell’intestino
viene assorbito la maggior parte, e solo 5 o 10 millimoli restano nell’intestino e
vengono eliminati tramite le feci. Circa 110mmol di sodio vengono trasportati nel
liquido extracellulare dove si uniscono al tutto quello che c’è. Qui il sodio è in
equilibrio con il liquido extracellulare perché continuamente entra nelle cellule
attraverso i canali di fuga e viene espulso continuamente tramite le pompe sodio-
potassio. Una piccola parte del sodio si perde tramite la pelle. Il sodio viene filtrato e
riassorbito nei reni e nell’urina se ne trovano circa 100millimoli al giorno.
Tanto sodio viene assunto, tanto sodio viene eliminato, poiché la quantità di sodio
presente nell’organismo deve rimanere costante.
Quasi tutto il sodio che viene filtrato deve essere riassorbito nel processo tubulare; 2/3 vengono riassorbiti
nel tubulo contorto prossimale.
È importante mantenere la concentrazione di sodio nel liquido extracellulare in quanto il sodio, essendo il
soluto del liquido extracellulare presente in quantità maggiore, è il maggior fattore dell’osmolarità del
liquido extracellulare; se cambia la concentrazione del sodio, cambia l’osmolarità del liquido extracellulare
e quindi si avviano dei processi di osmosi tra intra e extracellulare: se il sodio diminuisce fuori, l’acqua
tende ad entrare nelle cellule che si gonfiano ed esplodono o si rovinano, se invece il sodio extracellulare
aumenta, l’acqua viene allontanata dalle cellule che si raggrinziscono.
Inoltre se cambia la concentrazione di sodio cambiano anche le caratteristiche elettriche della membrana e
la sua polarizzazione.
Il mantenimento della concentrazione è importante inoltre perché permette di dar vita a tutti quei
meccanismi di trasporto attivo secondario che si basano proprio sul gradiente di concentrazione di sodio
transmembrana.
La stessa quantità di sodio che si ingerisce con la dieta, la si deve espellere (tramite soprattutto l’urina ma
anche tramite la sudorazione). Per cui si può magiare salatissimo oppure non mangiare un giorno, ma la
quantità di sodio presente nel corpo non cambia: il rene si adegua per espellere circa la stessa quantità di
sodio che viene assunta.
Il nefrone interviene nei suoi diversi tubuli per eliminare la giusta quantità di sodio.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
È già stato descritto sopra come avviene la filtrazione del sodio nel tubulo
contorto prossimale: la pompa sodio-potassio della membrana
basolaterale, determina l’entrata del sodio dalla membrana apicale
attraverso numerosi cotrasporti, canali ionici eccetera.
Nell’ansa di Henle (braccio discendente e la parte sottile del braccio ascendente) non ci sono grandi
cambiamenti; ci sono dei riassorbimenti passivi di sodio, per gradiente elettrochimico (di concentrazione e
del potenziale elettrico).
Nel tratto spesso del braccio ascendente dell’ansa di ha un imponente
riassorbimento di sodio. Questo avviene per la presenza di una pompa
sodio-potassio nella membrana basolaterale. Questo epitelio però a
differenza di quello del tubulo contorto prossimale è assolutamente
impermeabile all’acqua, quindi il riassorbimento di sodio e degli altri
soluti non può essere seguito dall’acqua. Il riassorbimento dei soluti va
ad aumentare la concentrazione dei soluti stessi fuori dal tubulo e a
diminuirla all’interno (in quanto non è stata eliminata una quantità di
acqua proporzionale alla quantità di soluti riassorbiti): il liquido tubulare
si diluisce. L’entrata di sodio dalla membrana apicale avviene con
meccanismi diversi rispetto a quelli del tubulo contorto prossimale. Entra una grande quantità di sodio per
gradiente di concentrazione in quanto la pompa sodio-potassio ha diminuito la concentrazione di sodio
all’interno della cellula. questo crea un gradiente tra liquido tubulare e citosol della cellula che porta il
sodio ad entrare. La principale via di entrata in questo tratto del nefrone è un cotrasportatore a tre che
trasporta un sodio, un potassio e 2 ioni cloruro. Se non c’è contemporaneamente la presenza di questi
quattro elementi, il trasporto non avviene. Il sodio (carica positiva) entra secondo gradiente elettrochimico,
portandosi dietro un potassio (carica positiva) e due ioni cloruro (2 cariche negative). questo fa si che il
sodio non si accumula nella cellula perché è già stato pompato all’esterno dalla pompa. Potassio e cloruro
invece si accumulano nella cellula ed escono della membrana basolaterale tramite canali ionici. Verranno
poi riassorbiti dal liquido extracellulare.
L’epitelio del braccio ascendente dell’ansa riassorbe una quantità enorme di sodio, potassio e cloruro.
Anche in questo epitelio inoltre è presente quel meccanismo di antiporto sodio-idrogeno, ma con
un’importanza minore.
Il riassorbimento del sodio nel tubulo contorto prossimale è simile a
quello del tratto spesso ascendente dell’ansa ma con un paio di
differenze. Si ha sempre una pompa sodio-potassio della membrana
basolaterale, ma anziché il cotrasportatore a 3 (sodio potassio e 2 ioni
cloro) si ha il costrasportatorre sodio-cloruro. Dal punto di vista elettrico
non ci sono grandi differenze perché in entrambi i casi entrano tante
cariche negative e tante cariche positive. La differenza importante è che
in questo tratto non viene riassorbito potassio. Inoltre, mentre l’epitelio
della parte spessa del tubulo ascendete era completamente
impermetabile all’acqua, l’epitelio del tubulo contorto distale presenta una permeabilità variabile all’acqua
a seconda del bisogno.
Nel dotto collettore si ha sempre la pompa sodio-potassio della
membrana basolatera e nella membrana apicale (o luminare) si hanno
canali del sodio e del potassio.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
RIASSORBIMENTO DEL CLORURO [Cl-]
Il cloruro tende sempre a riassorbire il sodio per equilibrio elettrostatico.
In particolare nel tubulo contorto prossimale ci sono due meccanismi di trasporto di
cloruro. Il primo meccanismo è detto para-cellulare in quanto non passa all’interno
delle cellule epiteliali, ma tra cellula e cellula, dal lume al liquido extracellulare (vedi
figura). Il passaggio del cloruro è dato dal fatto che sono stati riassorbiti degli ioni
sodio, che hanno formato una positività nel liquido extracellulare che attrae gli ioni
cloruro.
Un’altra parte passa per via transcellulare (passa all’interno della cellula epiteliale)
attraverso meccanismi di antiporto, e attraverso canali ionici sia nella membrana
apicale che in quella basolaterale.
Nel tratto spesso dell’ansa, il meccanismo fondamentale è il cotrasportatore a 3 (sodio, potassio, 2 cloruro).
Il cloruro accumulato all’interno della cellula uscirà per diffusione dalla membrana basolaterale attraverso
del trasportatori.
Nel tubulo contorto distale si ha il cotrasportatore sodio-cloruro attivato precedentemente dalla pompa
sodio-potassio della membrana basolaterale.
Nel dotto collettore si hanno diversi meccanismi, sia paracellulari che transcellulari, di cotrasporto e di
canali.
RIASSORBIMENTO DEL POTASSIO [K+]
Il potassio che si assume con la dieta è solitamente minore rispetto
al sodio. Ciò è abbastanza prevedibile, perché è quello che si
introduce con il cibo, e un po’ con l’acqua. Non si aggiunge potassio
solitamente, a meno che non si faccia una dieta mirata. Si assumono
solitamente circa 70 o 80 millimoli di potassio al giorno.
Una piccola parte del potassio che introduciamo con la dieta viene
eliminata con le feci, ma la maggior parte (circa 60millimoli al
giorno) viene riassoribita e va ad arricchire la quantità totale del
potassio nel liquido extracellulare; è in equilibrio con gli ambienti
intracellulari ricchi di potassio dei vari tessuti. La quantità di liquido
nello spazio extracellualre è quella che viene filtrata nel glomerulo
ed è quella che viene anche riassorbita. Venogno filtrati circa
800millimoli al giorno di potassio e riassorbiti circa 800millimoli al
giorno.
Per il potassio però, a differenza del sodio, si ha anche il
meccanismo di secrezione nel rene.
Si introducono 60 milli moli la giorno, quindi devono essere eliminati 60 millimoli. Se ne filtriamo 800 e ne
assorbiamo 800 millimoli, i 60 millimoli che dobbiamo eliminare escono dal processo di filtrazione. Il
processo di filtrazione lungo i tubuli è un’aggiunta rispetto al filtrato. Oltre ad essere riassorbito viene
aggiunta una quantità variabile di potassio a seconda delle esigenze, cioè di quanto ne è stato introdotto
nella dieta. In condizioni non fisiologiche però potrei avere una botta di potassio in eccesso: per esempio
quando si hanno delle lesioni cellulari. In questo caso il potassio che è solitamente accumulato nei liquidi
inracellulari delle cellule, a seguito di una lesione ampia viene rilascaito dalle cellule e va a finire nel lquido
extracellulare. Ciò non va bene in quanto se aumenta il potassio extracellualre il potenziale di membrana si
depolarizza perché diminuisce il numero di cariche positive che escono e che si spalmano sulla faccia
esterna della membrana.
Se cambio la concentrazione di uno ione, cosa succede alla diffusione di quello ione attraverso la
memebrana? Il potassio esce spontaneamente perché è più concentrato all’interno della cellula; questo
porta cariche positive all’esterno e genera il potenziale di membrana. Se aumento la concentrazone di
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
potassio intracellulare, la voglia del potassio di uscire dalla cellula diminuisce perché diminuisce il gradiente
di concentrazione, quindi diminuiscono le cariche positive che si accumulano all’esterno.
In base alla quantità di potassio che viene assunto, nel nefrone si attivano
meccanismi differenti:
- In condizioni di bassa assunzione di potassio il nefrone deve cercare di
trattenere il potassio presente nel filtrato e non lasciarlo nell’urina.
Consideriamo 100% il potassio che viene filtrato dal glomerulo, cioè è la
quantità totale giornaliera di potassio presente nel filtrato.
Nel tubulo contorto prossimale, ne viene riassorbito circa l’80%; nella parte
sottile dell’ansa di Henle non viene praticamente assorbito; nel tratto
spesso del braccio ascendente ne viene assorbito circa il 10%; il 10%
restante ne viene riassorbito in parte nel tubulo contorto distale, e il resto
nel dotto collettore. In questo modo alla fine più o meno tutto il potassio
filtrato è stato riassorbito.
- In condizioni di grandi assunzioni di potassio, o in condizioni non fisiologiche
(esempio malattia traumatica sopra citata), il potassio è presente in
concentrazioni maggiori.
Il riassorbimento resta uguale sia nel tubulo contorto prossimale, sia nella
parte sottile e spessa dell’ansa di Henle (fino a qui si ha un riassorbimento del
90% del totale); nel tubulo contorto distale si attiva invece un meccanismo di
secrezione del potassio anche molto intenso che può determinare una
secrezione importante anche uguale o maggiore rispetto alla quantità che è
stata filtrata.
Nel dotto collettore ne viene poi riassorbita una buona parte.
È un meccanismo di sicurezza che permette di eliminare il potassio che si è
accumulato in più nello spazio extracellulare.
RIASSORBIMENTO DELL’ACQUA
Il sodio viene riassorbito nei vari tratti con vari meccanismo, e a questa grande
quantità di sodio vanno dietro alcuni soluti, ma soprattutto l’acqua. L’acqua segue il
sodio per mantenere un equilibrio osmotico; l’uscita del sodio richiama l’acqua
all’esterno.
L’acqua per uscire dal tubulo passa passivamente per una via paracellulare (giunzioni
tra cellula e cellula), e attraverso la via transcellulare: nella membrana apicale e
basolaterale ci sono delle proteine, le acquaporine, che sono dei canali di acqua. Le
acquaporine sono una famiglia di proteine di membrna, transmembrana, che
formano un foro all’interno e che permettono il passaggio di acqua, sempre in modo
passivo (secondo un processo di osmosi).
Nel tratto spesso dell’ansa di Henle l’acqua non passa perché le giunzioni tra lecellule sono molto strette
(non permettono il passaggio di acqua per via paracellulare) e perché le cellule apiteliali sono prive di
acquaporine.
Nel tubulo contorto distale e nel dotto collettore la permeabilità dell’acqua è variabile: l’acqua non può
passare per via paracellulare ma può passare per via transcellulare, a patto che ci siano le acquaporine.
In questi tratti del tubulo la presenza di acquaporine è controllata: possono eserci come non esserci.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il riassorbimento dell’ACQUA e del SODIO nel tubulo contorto prossimane è accoppiato (vanno di pari
passo). Questi riassorbimenti sono regolati da diversi meccanismi di regolazione:
- Uno di questi è l’angiotensina 2 che viene prodotta nel sangue ad opera prima dall’enzima renina
(secreto dall’apparato iuxtaglomerulare) e poi dall’apparato ACE che si trova nella membrana dei
capillari capillari. Essa viene prodotta in condizioni di abbassamento della concentrazione di sodio
nel tubulo contorto distale. L’angiotensina 2 è un vasocostrittore che ha anche una funzione sul
riassorbimento del sodio, aumentandolo, agendo sul tubulo contorto prossimale, sul segmento
spesso dell’ansa di Henle e sul dotto collettore. Aumenta l’efficienza dei trasportatori del sodio.
- L’aldosterone è un ormone di grandissima importanza, prodotto dalla corticale surrenale, sotto
controllo dell’ngiotensina 2. L’aldosterone aumenta il riassorbimento di sodio agendo sul tratto
spesso dell’ansa di Henle e sul dotto collettore.
- Il peptide antidiuretico atriale è un ormone prodotto dal miocardio atriale quando aumenta la
volemia. Esso diminuisce il riassorbimento di sodio, e quindi anche di acqua, agendo sul dotto
collettore. Diminuendo il riassorbimento di sodio e di acqua fa perdere una quantitò maggiore di
urina e quindi diminuisce il volume ematico.
- L’urodilatina è un polipeptide podotto dalla parete del tubulo contorto distale e dal dotto collettore
in condizioni di pressione arteriosa aumentata. Essa diminuisce il riassorbimento di sodio e di
acqua. Ha la stessa funzione dell’ormone antidiuretico atriale, solo che quest’ultimo agisce a livello
generale passando nel sangue e ha anche altri effetti, mentr l’urodilatina ha un’azione locale
all’interno del rene.
- La guanilina è un polipeptide. È simpatica perché ha un meccanismo di feedforward per il sodio in
quanto viene liberata nel sangue (quindi è un ormone) dal colon quando viene introdotto un’alta
quantità di sodio cloruro. L’ormone agisce sul tubulo contorto prossimale diminuendo il
riassorbimento di sodio.
- L’adrenalina e la noradrenalina secrete in condizioni di stress rispettivamente dalla midollare
surrenale e dalle terminazioni norvose del sistema nervoso ortosimpatico, aumentano il
riassorbimento di sodio e di acqua agendo sull’ansa di Henle e sul dotto collettore.
30/04/2020
Nei tratti sottili dell’ansa di Henle viene riassorbita un po’ di acqua perché questi tratti si trovano nella
midollare, che presenta un ambiente extracellulare iperosmotico. Quindi anche se in questi tratti non c’è un
riassorbimento di sodio, l’ambiente iperosmotico permette il riassorbimento dell’acqua del 15%.
L’80% o più di acqua (che corrisponde a più di 130 litri al giorno) viene riassorbita prevalentemente nel
tubulo contorto prossimale e in parte minore, ma comunque importante, nell’ansa di Henle (parte sottile).
Dal punto di vista omeostatico questi riassorbimenti non vengono fatti in funzione dello stato di idratazione
o di disidratazione dell’organismo: l’acqua viene riassorbita a prescindere dallo stato di idratazione. L’acqua
riassorbita in funzione del bisogno del corpo di riassorbire sodio. Il riassorbimento non serve per regolare
l’omeostasi idrica, ma quella del sodio, e anche del potassio e del cloro.
Una volta riassorbita la maggior parte della quatità di acqua, il riassorbimento del sodio e dell’acqua e delle
loro omeostasi si dissociano. Quindi il 15 o 20% di acqua che resta da assorbire per arrivare ad 1litro e
mezzo di urina complessivo al giorno, deve essere mirato sul problema dell’omeostasi dell’acqua: è
regolato a seconda dell’idratazione giornaliera (data dalla quantità di acqua bevuta, dalla sudorazione ecc.).
Il riassorbimento obbligatorio dell’acqua corrisponde dunque alla prima parte riassorbita (85%), perché
corrisponde alla quantità di acqua che viene sempre riassorbita e non è regolato dalla disponibilità di acqua
del corpo; il riassorbimento facoltativo di acqua è quello che può essere effettuato o no, a seconda del
bisogno idrico di quel momento.
L’aldosterone è un ormone steroideo, quinidi derivato dal colesterolo (Il colesterolo ha una struttura di
base di 4 anelli di carboni). All’interno delle nostre cellule si formam l’aldosterone a partire dalla molecola
di colesterole, mantenendone le caratteristiche. L’aldosterone, derivando dal colesterolo è una molecola
lipoolubile (ha una prevalenza di legami nonpolari). La molecola quindi può attraversare facilmente il
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
doppio strato fosfolipidico e può avere un recettore
intracellulare. L’aldosterone è secreto dalle ghiandole
surrenali. Le ghiandole surrenali sono costituite da una parte
corticale e una midollare; le due parti sono completamente
diverse dal punto di vista istologico, fisiologico (secernono
ormoni diversi), embriologico. La parte corticale è divisa in 3
parti: glomerulare, fascicolata e reticolata. La parte esterna,
cioè quella glomerulare è quella che secerne l’aldosterone.
L’aldosterone essendo un ormone steroideo liposolubile, ha
dei recettori intracellulari. Entra all’interno della cellula del
tubulo, si unisce al suo recettore. Recettore e aldosterone
insieme si dimerizzano e poi entrano all’intenro del nucleo, dove il recettore si attacca a sequenze
specifiche di nucleotidi del DNA e attivano la sintesi di geni specifici. I geni e le proteine che vengono
sintetizzate per aumentare il riassorbimento di sodio sono la pompa sodio potassio; l’aldosterone
promuove la sintesi della pompa sodio-potassio. Il complesso aldosterone-recettore stimola la trascrizione
dei geni della pompa sodio potassio, dei canali del sodio e anche dei canali del potassio (che vanno a
collocalsi sulla membrana apicale). L’aldosterone promuove, oltre al riassorbimento di sodio, anche
l’eliminazione del potassio. Il potassio, che è entrato nella cellula tramite la pompa sodio-potassio, se trova
i canali sulla membrana apicale, allora effettivamente va ad aggiungersi al filtrato.
La presenza di aldosterone quindi aumenta il riassorbimento di sodio, a cui va dietro il cloro, e quindi anche
il riassorbimento dell’acqua. Aumenta la quantità di acqua riassorbita che va a finire nel sangue, quindi
aumentano il volume ematico e la pressione del sangue arteriosa e venosa. L’ormone è un ormone
ipertensivo, cioè che aumenta a pressione arteriosa.
Anche il cortisolo è un ormone di risposta allo stress e ha un po’ la stessa funzione dell’aldosterone, nel
senso che si può legare (con bassa affinità) allo stesso recettore dell’aldosterone.
Anche il colesterolo può andare ad aumentare il riassorbimento del sodio ed essere ipertensvo. Però
normalmente nelle cellule è attivo un enzima che prende il cortisolo e lo trasfomra in un’altra molecola
inattiva sui recettori dell’aldosterone. È vero che ogni recettore è specifico per un recettore, ma a volte non
in modo assuloto: un ormone ha i suoi effetti, ma può avere anche gli effetti di un altro ormone. Gli effetti
dell’aldosterone possono essere dati anche dal cortisolo. C’è un enzima che trasforma costantemente il
cortisolo in una molecola che invece non è capace di interagire con il recettore dell’aldosterone. Il
componente attivo delle liquerizia (acido glicirretico) inibisce specificamente questo enzima (quello che
trasforma il cortisolo in una molecola non il grado di agire sui recettori dell’aldosterone). La liquirizia fa si
che il cortisolo vada ad agire sui recettori dell’aldosterono e che quindi abbia il suo effetto ipertensivo
dovuto al maggior riassorbimento di acqua. La liquirizia ha questo effetto soprattutto in condizioni di stress,
poiché abbiamo molto cortisolo nel corpo. Quindi se mangiamo liquirizia e facciamo attività fisica intensa, la
pressione del sangue si alza.
RIASSORBIMENTO FACOLTATIVO DELL’ACQUA
Il volume di urina può variare molto: da mezzo litro a diversi litri al giorno.
Parallelamente al volume totale di uina prodotta, cambia il colore di urina: quando se ne fa tanta è molto
chiara, quando invece se ne fa poca diventa giallo intenso, anche marroncino. Ciò è indice della
concentrazione di soluti all’interno dell’urina. Ciò che cambia non è la quantità di soluti, ma la quantità
dell’acqua in cui si sciolgono i soluti: la quantità di soluti è sempre la stessa. Il colore dell’urina è dato da
molecole che derivano da componenti della bile. La bile (colorata con i pigmenti biliari) è prodotta dalla
cistifellea del fegato, viene riversata nell’intestino e assorbita dalle mucose intestinali. La si ritrova poi
nell’urina.
L’acqua viene riassorbita nei tubuli, sempre per osmosi (senza trasporto attivo).
Ma come si creano le condizioni di differenza osmotica tra il tubulo e il liqudo circostante in modo da
permettere il riassorbimento regolato, in base allo stato di idratazione? Ciò avviene grazie a due condizioni:
-la creazione di un ambiente iperosmotico nella midollare del rene. In questo modo non si crea una
situazione di equilibrio, infatti tutti i soluti presenti nell’ambiente iperosmotico tenderebbero a disperdersi
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
in tutto l’organismo. Si ha invece una diminuzione locale di entropia (i liquidi restano concentrati nella
midollare, non si disperdono) che richiede un dispensio di energia. Per creare e mantenere l’iperosmolarità
della midollare surrenare si necessita di spendenre energia metabolica, quindi ATP. L’ambiente inoltre
diventa sempre più iperosmotico andando in profondità nella midollare del surrene.
-il tubulo che passa attraverso l’ambiente iperosmotico deve avere una prmeabilità all’acqua variabile; deve
essere modulabile a seconda delle esigenze e delle condizioni di idratazione.
Per osmolarità si intende il numero di moli di particelle di qualsiasi tipo che sono presenti in un litro di
soluzione, o di solvente (se si parla di osmolalità). L’osmolarità è la misura del numero di particelle nel
soluto. È la misura con cui una soluzione tende a richiamare acqua a sé per osmosi.
Nell’immagine a fianco si può vedere un grafico in cui è rappresentato
il cambiamento dell’osmolarità in base alle zone del nefrone.
La curva blu indica l’osmolarità in condizioni di iperidratazione.
La curva rossa indica l’osmolarità in condizioni di disidratazione.
“1” indica il valore di osmolarità del sangue, quindi anche del filtrato
(in quanto i soluti del sangue sono uguali a quelli del filtrato, ad
eccezione per le proteine). In realtà questo 1 corrisponde ad un valore
di circa 300 milliosmolare per litro.
Lungo il corso del tubulo contorto prossimale l’osmolarità non cambia
perché viene riassorbito sodio e proporzionalmente acqua.
Nell’ansa di Henle, nella parte discendente, l’osmolarità aumenta fino
ad un valore di circa 1200 milliosmolari e poi diminuisce
drasticamente (soprattutto nel tratto spesso) fino ad arrivare ad un
valore inferiore a quello di partenza, circa a 100milliosmolari; per cui alla fine dell’ansa di Henle si ha un
liquido più diluito di quello iniziale del filtrato. Alla fine dell’ansa di Henle si arriva allo stesso valore sia in
condizioni di idratazione, sia in condizioni di disidratazione, ma attraverso due percorsi diversi: in caso di
iperidratazione c’è un aumento di osmolarità piccolo; nel caso di disidratazione l’aumento di osmolarità è
molto più grande. Alla fine del’ansa di Henle in entrambi casi si raggiunge lo stesso valore.
Nel tubulo contorto distale in condizioni di iperidratazione l’osmolrità continua a diminuire anche lungo il
dotto collettore, fino ad arrivare anche a valori estremi di 1/6 o 1/7 rispetto al valore dell’osmolarità
iniziale: è possibile produrre un’urina che è più diluita rispetto al filtrato di 6 o 7 volte (15litri in un giorno).
In caso di disidratazione, dopo l’ansa di Henle, l’osmolarità torna ad aumentare nel tubulo contorto distale.
Continua poi ad aumentare ancora di più nel percorso del dotto collettore potendo arrivare teoricamente
anche a valori di 3 o 4 volte più grandi rispetto a quello iniziale (fino anche a 1000 osmomolari).
I valori descritti dalle curve sono valori estremi, insuperabili, oltre i quali non si può andare. Normalmente si
hanno valori intermedi, tali per cui il valore di osmolarità dell’acqua è di poco superiore a quella del plasma.
[L’acqua di mare ha una concentrazione di cloruro di sodio più alto rispetto a quello dell’urina concentrata a
valori massimi. Nel caso in cui bevessimo acqua di mare, per eliminare quella enorme quantità di cloruro di
sodio assunto, dobbiamo eliminare anche una grande quantità di acqua. Per cui in caso di necessità bere
acqua di mare fa aumentare la dispersione di liquidi. Non bere acqua di mare! ]
In fiura sotto è rappresentato un altro schema riassuntivo di tutti i valori dell’osmolarità dell’acuqa lungo il
tubilo.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Per comprendere come si verificano tutte queste variazioni di osmolarità, le prime cose da tener presenti
sono le permeabilità e i trasporti che avvengono in alcuni tratti del tubulo.
Permeabilità= la membrana si la scia attraversare secondo gradiente elettrochimico. (in tabello è indicato
con il simbolo +)
Trasporto attivo= la membrana prende un soluto da una parte e forzatamente lo sposta dall’altra parte. È
indimendente dal gradiente di concentrazione. in tabella è indicato con il simbolo T.
ADH= ormone antidiuretico; in figura indica una permeabilità variabile a seconda o meno della presenza di
questo ormone.
H2O:
Il tubulo contorto prossimale è molto permeabile
all’acqua, anche l’ansa di Henle discendente; la parte
ascendente sottole dell’ansa di Henle è
sostanzialmente impermeabile all’acqua; il braccio
spesso dell’ansa è assolutamente impermeabile
all’acqua. Il tubulo contorto distale e il dotto
collettore sono permeabili solo in presenza
dell’ormone antidiuretico.
NA+: il tubulo contorto prossimale ha un meccanismo intenso di trasporto attivo. Nella parte idscendente
dell’ansa quasi non c’è trasporto attivo, ma è permeabile; la parte ascendente sottile è permeabile, ma
senza trasoporto attivo di sodio; nel tratto spesso dell’ansa c’è un trasporto attivo intensissimo (pompa
sodio-potassio accoppiata a un cotrasportatore a 3 di sodio potassio e due cloruri); la stessa cosa la si
ritrova nel tubulo contorto distale e nel dotto collettore.
Le stesse cose del sodio le si ritrovano anche nel cloruro.
UREA: L’urea è la molecola che viene formata nel fegato grazie al ciclo metabolico dell’urea. Essa per il
passaggio attraverso le cellule del tubulo richiede un trasportatore passivo. Se c’è il trasportatore può
passare per diffusione facilitata, altrimenti non passa. Di urea ne viene riassorbita poca nel tubulo contorto
prossimale; nell’ansa di Henle sottile, la membrana è permeabile;nel dotto collettore, in particolare
nell’ultima più profonda che va nellamidollare, l’urea è dipendente dall’ormone antidiudetico. Se non c’è
l’ormone l’urea è intrappolata nel dotto collettore; se c’è l’ormone l’urea esce per gradiente di
concentrazione.
Come si forma l’ambiente iperosmotico nella midollare? Qual è il meccanismo con lui l’ATP viene utilizzata
per concentrare i soluti nella midollare del rene tanto da avere un’osmolarità circa 4 volte superiore rispetto
a quella normale presente in tutte le altre parti del corpo?
Nel braccio spesso dell’ansa di Henle c’è il motore che mette in moto tutta una serie di eventi a cascata che
determineranno poi un’iperosmolarità sempre più grande a mano a mano che si va in profondità nella
midollare.
Nel tratto spesso del’ansa la pompa sodio-potassio riassorbe una grande quantità di sodio e sulla
membrana apicale è presnte il cotrasportatore a 3 (sodio potassio e 2 clori). In questa parte dell’ansa viene
riassorbita un grande quantità i sodio cloruro e potassio che vanno nel liquido extracellulare della
midollare. Ciò aumenta l’osmolarità della midollare. Il filtrato che rimane nel tubulo è stato diluito in
quanto sono stati tolti dei soluti e in quanto non è avvenuto passaggio di acqua poiché la membrana è
assolutamente impermeabile. Il liquido diluito ora presenta un’osmolarita di circa 1/3 rispetto a quella
insiziale (passa a 100milliosmolari) ed entra nel tubulo contorto distale.
Nel tubulo contorto distale la permeabilità all’acqua è variabile a seconda o meno dell’ADH.
Se c’è l’ormone: il tubulo contorto distale sta nella corticale del rene, che non ha subito il processo di
concentrzione dovuto ai trasporti che avvengono nel tratto spesso, quindi ha un’osmolarità normale di
300milliormolari. Per cui essendoci all’interno una osmolarità di 100milliosmolari e all’esterno di
300milliosmolari, l’acqua esce per gradiente di concentrazione (va da un ambiente più diluito, ad un
ambiente più concentrato per osmosi). Se l’acqua esce, il liquido nel tubulo contorto distale si concentra e
torna sostanziaemente ai valori di osmolarità normali, 300milliosmolari.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Per cui è stata riassorbita dell’acqua e nel tratto spesso sono stati riassorbiti dei soluti, ma c’è stata una
separazione tra riassorbimento di soluti e riassorbimento di acqua.
Questo liquido sbocca nel dotto collettore a 300milliosmolari. Il dotto collettore scende nella midollare,
dove c’è un ambiente iperosmotico. L’ambiente extracellulare della midollare è iperosmotico perché
precendentemente ci è stato buttato tanto sodio, cloruro e potassio. Se la parete del dotto collettore è
permeabile all’acqua (quindi se c’è la presenza dell’ormone) l’acqua viene richiamata all’esterno dalla
differenza di osmolarità. Se l’acqua esce, i vari soluti che si trovano all’interno del tubulo, si concentrano. In
realtà la pompa sodio potassio la si trova anche nel tubulo contorto prossimale e nel dotto collettore, per
cui questi soluti continuano ad uscire in prte, ma l’unico soluto che non può uscire è l’urea. Essa può uscire
solo nella parte più profonda del dotto collettore, a patto che ci sia l’ormone ADH. Se c’è l’ormone l’urea
esce dalla parte finale del dotto collettore per gradiente di concentrazione e va a finire nel liquido
extracellulare della parte profonda della midollare. Questo va ad aumentare ancora di più l’osmolarità della
parte profonda della midollare, che era già stata aumentata. A questo punto l’urea diventa più concentrata
nella parte profonda della midollare rispetto alle parti sottili delle anse. Poiché la parte sottile dell’ansa è
permeabile all’urea, l’urea per gradiente di concentrazione rientra all’interno del tubulo a livello della parte
sottile dell’ansa. A questo punto l’urea viene trascinata e ricircola nel tubulo contorto distale e nel dotto
collettore. La concentrazione di urea aumenta ancora, non solo perché è uscita una quantità d’acqua, ma
anche perché l’urea è stata riciclata. Questo giro del’urea che entra ed esce determina un aumento sempre
più grande della concentrazione di urea nel liquido extracellulare della midollare.
Si è creato quindi nella midollare un ambiente iperosmotico: nella parte più superficiale della midollare è
dovuto principalmente a cloruro sodio e potassio che vengon fuori dal tratto spesso; nella parte più
profonda è dovuto principalmente all’urea che esce dal dotto collettore.
Perché più si arriva in profondità e più i valori di osmolarità aumentano?
Nell’ansa di Henle avvengono due tipi di processi:
- il liquido scorre dal braccio discendente a quello ascendente
- dal tratto spesso braccio ascendente vengono pompati fuori soluti (sodio cloruro e potassio) senza
acqua, che venogno accumulati nel liquido extracellualre.
Questi soluti però non si fermano lì perché nel tratto discendente dell’ansa, questi soluti che hanno
aumentato l’osmolarità del liquido extracellulare tirano fuori l’acqua (infatti il tratto discendente è
permeabile all’acqua) e possono anche rientrare nel braccio discendente. Tirano fuori l’acqua fino a
far diventare l’osmolarità del braccio discendente uguale a quella del liquido extracellare
circostante. La potenza del meccanismo di estrazione dei soluti nel tratto spesso è tale che si può
stabilire una differenza di osmolarità di 200milliosmoli per litro.
Immaginiamo con la fantasia una condizione in cui questi processi non sono
ancora iniziati: ci sarebbe all’interno dell’ansa un liquido di 300 milliosmolari,
uguale a quello dell filtrato iniziale e come al’ambiente extracellulare. Quando i
soluti escono dal tratto spesso ascendente, si va a creare una differenza di
osmolarità di 200milliosmolari; per cui nel tratto ascendente ci sarà un valore di
200, mentre il liquido extracellulare avrà un valore di 400. Anche il braccio
ascendente tende ad assumere valori simili a quelli del liquido extracellulare,
per cui avrà un valore anch’esso di 400 milliosmolare.
Il liquido scorre all’interno dell’ansa. Quindi il valore di 300 che si trovana nel
liquido contorto prossimale scorre ed entra nel braccio discendente dell’ansa.
Per cui all’inizio del braccio discendente non c’è un valore di 400, ma un valore
di 300milliosmolare a causa dello scorrimento del liquido. Inoltre i 400
milliormolari che si trovano in fondo al braccio discendente dell’ansa,
scorrendo, arrivano nella parte più profonda del braccio ascendente.
Ripetuto questo porcesso “n” volte abbiamo una situazione reale.
Si parte quindi in una condizione in cui all’interno del tratto spesso del tubulo
ascendente si ha un valore di 200 e nelo spazio extracellulare c’è un valore di
300 milliosmolari (valore che si è omologato al valore del liquido che esce dal
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
tubulo contorto prossimale). L’uscita dei soluti dalla parte spessa del tubulo ascendente crea una differenza
di 200 milliosmolari tale per cui nel tubulo spesso ascendente si crea un valore di 150; nello spazio
extracellulare si crea un valore di 350. A seguito poi dell’uscita di acqua dal tratto discendente, anche una
parte del tratto discendente prende lo stesso valore dello spazio extracellulare (350).
Nella parte più profonda dell’ansan che non parte né da 300 come all’inizio, né da 200 com’era diventato,
ma da 400 a causa dello scorrimento. Se il riassorbimento stabilisce una differenza di 200 i valori
diventeranno 300 nella parte finale del tratto discendente e 500 nella parte iniziale del tratto ascendente.
Ripetendo ancora “n” volte questo processo ci si ritrova un’altra condizione ancora in cui si ha nella parte
profonda dell’ansa un valore di 600.
Si può dunque continuare a ripetere il procedimento per numerose volte, tenendo presente 3 cose:
1- I meccanismi di trasporto del braccio ascendente sono capaci di creare una differenza di osmolarità
di 200.
2- Il braccio ascendente dell’ansa è così permeabile all’acqua che si omogenizza subito con il liquido
extracellulare circostante.
3- Il liquido scorre dal discendente all’ascendente.
Il fatto che il processo si ripeta a vari livelli, determina una sempre maggiore osmolarità a mano a mano che
si scende nella midollare. Questo permette di tirar fuori sempre più acqua dal dotto collettore: nella
midllare si avrà un’osmolarità sempre più alta e quindi si tira fuori sempre più acqua da dotto collettore che
passa nella midollare.
VARIABILITA’ DELLA PERMEABILITA’ DELL’ACQUA
Se la permeabilità dell’acqua del tubulo contorto distale e del dotto collettore fosse zero, cioè se non ci
fosse l’ormone antidiuretico, l’osmolarità del liquido ottenuto nel tutbulo rimarrebbe quella che esce
dall’ansa di Henle, di 100milliosmolari. L’urina finale sarebbe un’urna fortemente diluita, diluita 3 volte
rispetto al valore iniziale e al valore del plasma. Senza l’ormone si arrivano a fare 15 litri di urina ad
osmolarità 100.
Se la permeabilità fosse massima sia nel tubulo contorto distale e nel dotto collettore, il liquido tubulare
perderebbe tanta acqua per osmosi, da far diventare l’osmolarità dell’urina, nella parte profonda della
midollare, uguale a quella del liquido circostante. Avrebbe un valore di 1200, cioè 4 volte più grande
rispetto al valore normale di osmolarità del plasma. Se c’è una grande quantità di ormone antidiuretico, si
fa mezzo litro di urina ad osmolarità 1200.
Il massimo di osmolarità a cui si può arrivare nella midollare è 1200, in qaunto il passaggio di acqua è
passivo e piò diventare uguale al valore dello spazio extracellulare.
Come fa l’ormone antidiuretico a rendere permeabile la parete del tubulo contorto distale e del tubulo
collettore?
Le membrane sono permeabili all’acqua se contengono le acquaporine. Queste proteine formano un foro
dove ci sono delle cariche che si affacciano sulla superficie del foro, in modo tale che le molecole di acqua
possono interagre con queste cariche.
In figura è rappresentata una cellula epiteliale del dotto collettore.
La membrana apicale è quella in alto; la membrana basolaterale è quella ai lati e in basso.
Nella membrana basolaterale le acquaporine sono sempre presenti.
Nella membrana apicale invece le acquaporine non ci sono. Queste
proteine si trovano nella membrana di vescicole (originate
dall’apparato di Golgi) che si trovano all’interno della cellula epiteliale
del dotto collettore. In particolare, nella membrana di queste vescicole
si trovano le acquaporine 2. Sulla membrana basolaterale c’è il
recettore dell’ormone atidiuretico, che è un ormone proteico, un
polipeptide. Esso non può attraversare la membrana, ma ha bisogno di
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
un recettore di membrana: questo è un recettore a 7 passi accoppiato a proteina G che attiva l’adenilato
ciclasi e che quindi lavora attraverso il meccanismo dell’AMP ciclico.
L’ormone antidiuretico (AVP in figura) si lega al suo recettore, che attiva la proteina G, che attiva l’adenilato
ciclasi, che trasforma l’ATP in AMP ciclico, che attiva la PKA. La PKA va a fosforilare delle proteine della
membrana delle vescicole che, una volta fosforilate, permettono il processo di esocitosi delle vescicole.
Quindi quando arriva l’antidiuretico questo determina la fosforilazione delle proteine della membrana della
vescicola e la vescicola si fonde con la membrana apicale della cellula eiteliale del dotto collettore.
Fondendosi per un processo di esocitosi, le sue proteine di membrana, in particolare le acqua porine vanno
a far parte della membrana apicale della cellula epiteliale del dotto. A questo punto la membrana che era
impermeabile all’acqua diventa permeabile all’acqua.
Ora l’acqua può entrare e una volta entrata può uscire tramite le acquaporine 4 che non sono dipendenti
dall’ormone antidiuretico, ma sono sempre presenti nella membrana basolaterale.
L’acqua esce dalla cellula attraverso le acquaporine4 proprio perché tutto questo avviene nel dotto
collettore che si trova immerso in nell’ambiente iperosmotico della midollare renale.
Quando c’è l’ormone antidiuretico l’acqua viene riassorbita nel tubulo contorto distale nel dotto collettore
e quindi l’urina diminuisce di volume e aumenta di concentrazione dei soluti per sottrazione di solvente.
DOVE E COME VIENE SECRETO L’ORMONE ANTIDIURETICO
L’ormone antidiuretico è un piccolo peptide, fatto da pochi residui amminoacidici. Esso viene secreto dalla
neuroipofisi. L’ipofisi è una ghiandola situata alla base del cervello ed è composta da due parti: neuroipofisi
(“neuro”= tessuto nervoso) e adenoipofici (“adeno”=ghiandola).
L’ipofisi proprio durante lo sviluppo embrionale deriva da due parti completamente
diverse (come la surrenale). La neuroipofisi deriva da un’estroflessione del primo
abbozzo di cervello ed è costituita proprio da tessuto nervoso; l’adenoipofisi deriva
da un gruppo ci cellule che originano da tutt’altra parte e che migrando
nell’embrione, si vanno ad attaccare alla neuroipofisi. L’origine e la funzione delle due
ghiandole sono completamente diverse, ma sono situate entrambe sotto l’ipotalamo
con cui hanno stretti rapporti.
La neuroipofisi è un tessuto nervoso ed è costituita all’interno da un fascio di assoni
di neuroni i cui corpi cellulari si trovano in due nuclei dell’ipotalamo (paraventricolare
e sopraottico).
L’ormone antidiuretico viene sintetizzato nei corpi cellulari dei neuroni. Da qui poi scende nella nauroipofisi
per via nervosa, attraverso gli assoni che giungono fino la neuroipofisi. Esistono dei meccanismi di trasporto
assonale (come delle rotaie di cotoscheletro) lungo cui corrono dei trasportatori (come dei trenini ) che
portano il materiale dal corpo cellulare fino all’estremità dell’assone.
Le terminazioni che si trovano nella neuroipofisi sono e funzionano come gli elementi presinaptici.
L’ormone antidiuretico si trova inscatolato all’interno di vescicole, che si comportano come vescicole
sinaptiche.
Quando i neuroni si ecitano, e arrivano all a soglia, parte un potenziale d’azione dal cono di emergenza
dell’assone che percorre tutto l’assone. Il potenziale d’azione arriva fino all’estremità e succede tutto quello
che succede in tutti gli elementi presinaptici: si aprono canali del calcio voltaggio dipendenti, entra calcio
che si lega alle proteine della membrana delle vescicole, determinandone l’esocitosi e quindi lo
sversamento del contenuto (che è in questo caso l’ormone antidiuretico) nel liquido extracellulare. Una
volta versato nel liquido extracellulare l’ormone entra nei capillari che se lo portano via.
L’ormone antidiuretico viene secreto nella neuroipofisi e viene sintetizzato nell’ipotalamo.
QUANDO VIENE SECRETO L’ORMONE ANTIDIURETICO
Ci sono diversi fattori che stimolano o inibiscono la secrezione dell’antidiuretico.
Primo di tutti l’iperosmolarita del liquido extracellulare dell’ipotalamo. L’ipotalamo viene irrorato dalla
circolazione sanguigna, per cui l’smolarità dell’ambiente extracellulare dell’ipotalamo è uguale a quella del
sangue. In uno stato di disdratazione il sangue, e quindi anche il liquido extracellulare, è iperosmotico.
Nell’ipotalamo ci sono dei neuroni che sono osmocettori, cioè si eccitano quando l’osmolarità
dell’ambiente aumenta. Questi neuroni sono mescolati insieme ai neuroni che producono l’ormone
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
antidiuretico e fanno sinapsi su questi neuroni. Quindi quando gli osmocettori si eccitano, attivano la
sinapsi sui neuroni che producono ormone antidiuretico e l’ormone viene rilasciato.
Se l’ambiente extracellulare diventa extracellulare la cellula si saggrinzisce perché perde acqua per osmosi.
Raggrinzendosi la cellula cambia forma, e se nella membrana ci sono dei canali cationici a controllo
meccanico vengono aperti. L’apertura dei canali determina l’entrata del sodio; si arriva alla soglia e parte il
potenziale d’azione che fa sinapsi sul neurone che secerne l’antidiuretico. L’antidiuretico va a permettere il
riassorbimento di acqua nel rene. Inoltre, trattenendo acqua e lasciando andare via i soluti con l’urina,
l’ambiente extracellulare dell’organismo di diluisce e si risolve il problema di essere in condizioni
iperosmotiche.
Altri fattori che controllano la secrezione dell’antidiuretico sono i volocettori atriali: aumenta il volume del
sangue, si distende l’atrio destro, vengono attivati i recettori di volume e questo, attraverso un via nervosa,
va all’ipotalamo e inibisce la secrezione di antidiuretico.
I barocettori arteriosi (situati nell’arco aortico e nella biforcazione delle carotidi): sono sensibili ad un
aumento della pressione arteriosa. La loro attivazione va ad inibire, tramite una via nervosa, la secrezione di
antidiuretico.
Questi due meccanismi indicano che se aumentano il volume e la pressione del sangue, il corpo, eliminando
acqua tramite l’urina, riporta i valori di volume e di pressione alla normalità.
Altri meccanismi sono i peptidi natriuretico atriale, la cui secrezione viene attivata da un aumento di
volume del sangue totale (quindi da una distensione dell’atrio destro) che inibisce la secrezione di
antidiuretico.
L’angiotensina 2 che è dipendente da una diminuzione della pressione arteriosa, aumenta la secrezione di
antidiuretico.
Altri fattori non fisiologici, che introduciamo sono l’alcol etilico e la nicotina. L’alcol etilico è un forte
inibitore della secrezione dell’ormone antidiuretico. La nicotina invece ha effetti opposti.
04/05/2020
È uno schema che riassume quasi tutto sul feedback della pressione arteriosa. La
pressione arteriosa del sangue è strettamente correlata con la concentrazione di
sodio e cloruro (perché richiamano acqua e fa aumentare il volume ematico).
attraverso tutti i vari passaggi dello schema si arriva alla compensazione delle
variazioni della pressione arteriosa.
CONTROLLO DEL pH
A livelli fisiologici il pH ha un valore di 7.39/7.40, mantenuto in modo molto rigido. Abbiamo già visto 2 dei
3 livelli di intervento per il mantenimento del pH:
Il livello ematico: i sistemi tampone, presenti nel sangue, permettono di assorbire gli ioni idrogeno quando
aumentano, e rilasciarli quando diminuiscono. Esempi di sistemi tampone sono il sistema acidocarbonico
bicarbonato; il sistema fosfato acido fosfato alcalino; il sistema dato dalle proteine plasmatiche.
Il livello della respirazione: la respirazione polmonare attraverso il cambiamento di ventilazione alveolare
può modificare la quantità e la velocità di eliminazione di CO2. Essendo la CO2 in relazione con il pH
attraverso le due reazione, essa può intervenire nella sua regolazione.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Il livello renale: è fondamentale rispetto agli altri: infatti il primo livello si può esaurire (i sistemi tampone
non sono infiniti) e il secondo livello determina la diminuzione degli ioni idrogeno ma anche degli ioni
bicarbonato, andando a diminuire l’efficienza del primo sistema tampone (acidocarbonico-bicarbonato). Ciò
vuol dire che è vero che con la respirazione si compensa la tendenza all’acidosi, ma ci si pone anche a rischi
di acidosi ulteriore, a causa della diminuzione del primo sistema di tampone. È un impoverimento della
riserva alcalina.
Il rene elimina ioni idrogeno fisicamente. Metre la respirazione elmina CO 2 e quindi indirettamente
diminuisce la concentrazione di ioni idrogeno perché si associano a bicoarbonato, il rene elimina
definitivamente gli ioni idrogeno dal corpo. Per la prima volta gli ioni idrogeno di troppo non vengono
nascosti dentro una molecola o dentro un sistema tampone, ma vengono eliminati tramite l’urina.
Inoltre il rene risolve il problema aperto dalla respirazione: oltre a buttar fuori gli ioni idrogeno, il rene
cattura, recupera, degli ioni bicarbonato, quindi torna a ricostituire quella riserva alcalina che si era
impoverita con l’intervento della respirazione.
Questo avviene in quasi tutta la lunghezza del nefrone acetto che nella parte sottile dell’ansa di Henle. Nella
cellula epiteliale del tubulo contorto prossimale (ma anche nel resto del nefrone) c’è un enzima, l’anidrasi
carbonica, che prende la CO2 + H2O e forma acido carbonico che poi si dissocia in parte in ioni idrogeno e
ioni bicarbonato. L’epitelio del tubulo renale è un epitelio polarizzato, per cui i trasportatori della
membrana apicale sono diversi da quelli della membrana basolaterale.
Nella membrna apicale c’è uno scambiatore sodio-idrogeno: il sodio che entra
all’interno della cellula fa uscire con un meccanismo di antiporto lo ione idrogeno
che si è formato all’interno della cellula epiteliale. Chiaramente è sempre il sodio
che gestisce la situazione: esso entra nella cellula grazie al gradiente di potenziale
elettrochimico (gradiente di concentrazione + gradiente di potenziale elettrico).
Lo scambiatore sodio-idrogeno, pompa ioni idrogeno nel liquido tubulare.
Gli ioni bicarbonato che si sono formati dalla dissociazione dell’acido carbonico,
hanno trasportatori solo nella membrana basolaterale. Questi trasportatori sono
diversi, tra cui il meccanismo di cotrasporto 1sodio e 3 bicarbonati. In questo
caso l’energia viene fornita dal bicarbonato che è ampiamente presente nella
cellula e che diffonde verso il liquido extracellulare per gradiente elettrochimico,
trasportandosi dietro il sodio. Il bicarbonato dal liquido extracellulare viene preso dal flusso ematico dei
capillari peritubulari e viene trasportato tramite il sangue in tutti i tessuti.
Lo ione idrogeno che è stato espulso, si può anche ricombinare con il bicarbonato (o unirsi ad altre
molecole), formando acido carbonico.
Nelle cellule epiteliali sono presenti diverse isoforme di anidrasi carbonica: una (di cui abbiamo gia parlato)
si trova all’interno del citosol della cellula; l’altra è un’anidrasi carbonica di membrana che lavora sul
versante del lume della membrana apicale. Questa anidrasi carbonica può prendere l’acido carbonico che si
è formato grazie all’unione dello ione idrogeno (che è stato espulso) con il bicarbonato tubulare (che si
trova già nel liquido tubulare), e si forma acido carbonico. L’anidrasi carbonica compie la reazione: da acido
carbonico ad anidride carbonica. L’anidride carbonica che entra nella cellula viene riconvertita in acido
carbonico reagendo con l’acqua (CO2+H2O=H2CO3 H2CO3= H+ + HCO3). A questo punto gli ioni idrogeno
vengono espulsi dalla cellula verso il liquido tutbulare e ricomincia il ciclo. Ad ogni “giro” si tirano dentro la
cellula 3 ioni bicarbonato che vengono poi inviati al liquido extracellulare e poi al sangue.
Tutto ciò avviene nel tubulo contorto prossimale. Il nostro corpo sempre tende all’acidosi, sia se stiamo
fermi e soprattutto se ci muoviamo: nel tubulo contorto prossimale si elimina quanto più prossibile il
problema dell’eccesso di idrogeno. l’aumento di bicarbonato porta ad un aumento di pH.
L’anidride carbonica all’interno della cellula del tubulo proviene in piccola parte dai capillari peritubulari
che per diffusione forniscono CO2 alla cellula tubulare; ma in massima parte la CO2 proviene dallo stesso
filtrato. Infatti è la quota fisicamente disciolta presente nel plasma del sangue venoso ma anche del sangue
arterioso. L’anidride carbonica diffonde facilmente attraverso le membrane, per cui entra nella cellula
tubulare.
Gli ioni idrogeno possono essere espulsi verso il lume del tubulo anche da un trasportatore sodio ione
ammonio. Questo è ancora più importante perché unisce ancora un altro vantaggio.
Lo ione ammonio (NH4+) deriva dalla ammoniaca (NH3) che si unisce ad uno ione idrogeno (NH 3+H+=NH4+).
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
L’ammoniaca si forma all’interno della cellula tubulare del tubulo prossimale a partire dalla glutammina che
viene deamminata (tolto un gruppo amminico) per dare l’acido glutammico (glutammato). L’ammoniaca si
unisce ad uno ione idrogeno formando NH4+. Lo ione ammonio viene espulso con un meccanismo di
antiporto sodio-ammonio. Il vantaggio ulteriore, oltre all’eliminazione di uno ione idrogeno e al recupero di
un bicarbonato, è l’eliminazione dell’ammoniaca. L’ammoniaca è fortemente tossica.
Nel tratto spesso dell’ansa di Henle ci sono la pompa, il cotrasportatore a 3, e anche gli
stessi meccanismi di trasporto di ioni idrogeno e bicarbonato, pre
senti nel tubulo contorto prossimale. L’anidrasi carbonica forma acido carbonico che si
dissocia; gli ioni idrogeno vengono trasportati nel lume del tubulo dal meccanismo di
antiporto sodio-idregeno (alimentato dalla pompa sodio potassio) e lo ione
bicarbonato viene espulso nel liquido extracellulare con quel meccanismo di antiporto
cloro-bicarbonato.
Nell’ultima parte del tubulo contorto distale e nel dotto collettore avvenegono le rifiniture: vengono definiti
quanti ioni idrogeno devono essere espulsi precisamente e quanti ioni bicarbonato devono essere
riassorbiti in base al pH del sangue e del liquido extracellulare dellìorganismo in quel preciso momento.
Tuto questo a sua volta dipende da un’nfinità di cosa: dal tasso metabolico, dalla capacità respiratoria, dalla
dieta, dall’efficienza dei sisitemi tampone.
Nel dotto collettore in particolare si hanno 3 tipi di cellule ndoteliali: le cellule principali, le cellule
intercalate di tipo A (alpha) e le cellule intercalate di tipo B (beta). Le cellule intercalate sono quelle di
numero maggiore.
Le cellule principali hanno funzioni molto simile a quelle del tubulo contorto prossimale e al tratto spesso
dell’ansa (l’anidrasi carbonica, produce ioni idrogeno che venogno inviati nel lume grazie al meccanismo di
antiporto con il sodio; nella membrana basolaterle ci sono i meccanismi di trasporto del bicarbonato).
Quindi tramite queste cellule si continua ad eliminare il grosso degli ioni idrogeno e a recuperare il
bicarbonato.
Le cellule intercalate di tipo A dal punto di vista metabolico sono simili a quelle vste precedentemente.
L’anidrasi carbonica prende la CO2, la fa reagire con l’acqua e forma ioni idrogeno e
bicarbonato. Il bicarbonato viene pompato dalla membrna baso laterale verso il liquido
extracellulare. Gli ioni idrogeno in parte vengono messi nel lume del tubulo da vari
meccanismi di antiporto. L’altra parte di ioni idrogeno sfruttano un meccanismo Di
trasporto attivo primario presente nella membrana plasmatica apicale della cellula
intercalata di tipo A. Questo meccanismo, essendo primario, consuma ATP, infatti la
proteina è un ATPasi e ha la funzione di trasportare solo ioni idrogeno contro gradiente
di concentrazione. Avviene contro gradiente di concentrazione in quanto già sono
avvenuti tanti trasporti di ioni idrogeno e il liquido tubulare si è già acidificato. Infatti
questo meccanismo avviene proprio grazie all’energia data all’idrolisi dell’ATP.
Fino a questo punto il trasporto di ioni idrogeno nel lume è avvenuto grazie all’entrata di sodio (è l’entrata
di sodio che formisce energia per l’eliminazione dello ione idrogeno).
A questo punto potrei avere bisogno di riassorbire ioni sodio oppure non avere più bisogno di riassorbirli
ma essere allo stesso tempo in acidosi, e avere quindi un ulteriore bisogno di eliminare ioni idrogeno. in
questo ultimo caso si deve rendere indipendente l’eliminazione ulteriore di ioni idrogeno dal
riassorbimento parallelo di ioni sodio. il meccanismo delle cellule intercalate permette di intervenire
sull’omeostasi degli ioni idrogeno indipendentemente dall’omeostasi degli ioni sodio (e di qualsiasi altra
omeostasi). È necessario avere ATP a sufficienza per eliminare ioni idrogeno.
Le cellule intercalate di tipo B sono delle cellule rovesciate, in cui il meccanismo di antiporto bicarbonato-
cloruro è posto sulla membrana apicale (non più su quella basolaterale) il meccanismo di trasporto attivo
primario è posto sulla membrana basolaterale (anziché in quella apicale). È tutto al rovescio.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
L’anidrasi carbonica prende la co2 e forma ione bicarbonato e ioni idrogeno: per cui si ha
un riassorbimento attivo di ioni idrogeno che venogno portati nel sangue e un
eliminazione di ione bicarbonato.
Il riultato di queste cellule è quello di acidificare il sangue e alcalinizzare l’urina.
Queste cellule saranno utili quando arrivati al tubulo contorto distale o al dotto
collettore sono stati eliminati troppi ioni idrogeno (quindi si è in una condizine di
alcalosi). Per esempio qunaod vado in alta montagna, cambio altitudine si va in carenza
di ossigeno perché l’aria è più rarefatta e si va in una situazione di ipossia che si traduce
in ipossiemia. Per compensare lo scarso ossigeno si aumenta la ventilazione, quindi si
elimina una quantità maggiore di CO2. Eliminando una quantità maggiore di CO2, pur
non essendo in acidosi (quindi non avendo una produzione maggiore di CO2), il corpo va in alcalori, cioè
aumenta il pH. In questa situazione le cellule intercalate di tipi B sono fondamentali, in quanto sono stati
eliminati troppi ioni idrogeno e il corpo è addirittura in alcalosi.
MECCANISMI DI CONTROLLO DELLE CELLULE
- pH del liquido extracellulare del rene: se sono in acidosi o in alcalosi, anche il liquido extracellulare
del rene diventa acido o basico. Lo stesso pH del liquido extracellulare va a modificare i meccanismi
delle cellule epiteliali del tubulo. Una diminuzione di pH auementa, nella membrana apicale,
l’aggiunta del meccanismo di antiporto sodio-ioni idrogeno.
- le cellule endoteliali dei capillari peritubulari e le cellule del tubulo contorto prossimale producono
l’endotelina che è una proteina che agisce aumentando l’efficienza del trasporto sodi idrogeno
nella membrana apicale e di bicarbonato nella membrana basolaterale. L’endotelina viene regolata
sulla base del pH del sangue che scorre nei capillari. Una variazione di pH determina la secrezione di
endotelina che agisce localmente sul rene, aumentando l’efficienza del rene nella eliminazione di
ioni idrogenoe recupero di bicarbonato
- cortisolo (ormone)
- aldosterone (ormone)
- angiotensina 2 (ormone)
Questi ultimi 3 ormoni vengon secreti sulla base di diversi fattori di controllo, tra cui il pH del sangue. Il pH
va a modificare la secrezione di questi ormoni che vanno ad agire sulle cellule intercalari A e B, modificando
la funzione relativa. Determinano l’eliminazione o il riassorbimento di ioni idrogeno.
Document shared on https://www.docsity.com/it/sbobine-funzione-renale-fisiologia/8059847/
Downloaded by: EnricoCusino (e.cusino@campus.uniurb.it)
Potrebbero piacerti anche
- Linfedema: Spunti pratici per la terapia multidisciplinareDa EverandLinfedema: Spunti pratici per la terapia multidisciplinareNessuna valutazione finora
- Sistema EscretoreDocumento17 pagineSistema Escretorewz79zb96h6Nessuna valutazione finora
- Zanzare in Appenino - I culicidi di alta quota in provincia di ModenaDa EverandZanzare in Appenino - I culicidi di alta quota in provincia di ModenaNessuna valutazione finora
- Il FegatoDocumento7 pagineIl FegatoNicola BombaceNessuna valutazione finora
- La terapia fisica vascolare - la prossima generazione della medicina?: La microcircolazione del sangue - Quello che tutti dovrebbero sapereDa EverandLa terapia fisica vascolare - la prossima generazione della medicina?: La microcircolazione del sangue - Quello che tutti dovrebbero sapereNessuna valutazione finora
- Apparato UrinarioDocumento17 pagineApparato UrinarioCate?Nessuna valutazione finora
- Ginnastica respiratoria - Purificazione - Salute - Forza - Energia mediante l'esercizio fisico e psichico della respirazioneDa EverandGinnastica respiratoria - Purificazione - Salute - Forza - Energia mediante l'esercizio fisico e psichico della respirazioneValutazione: 4 su 5 stelle4/5 (1)
- Apparato UrinarioDocumento20 pagineApparato UrinarioBruce WiiNessuna valutazione finora
- Nefrologia e UrologiaDocumento332 pagineNefrologia e UrologiaMatteoDiNassoNessuna valutazione finora
- Dissezione Dei ReniDocumento16 pagineDissezione Dei Renivincenzo fasanellaNessuna valutazione finora
- Appunti UrologiaDocumento29 pagineAppunti UrologiaMatteo Gidgio MeloniNessuna valutazione finora
- Apparato UrinarioDocumento3 pagineApparato UrinarioputtanaNessuna valutazione finora
- Apparato Urinario RiassuntoDocumento4 pagineApparato Urinario RiassuntoMastromauroNessuna valutazione finora
- Patologia Clinica CompletaDocumento75 paginePatologia Clinica CompletaMatteo TomaselliNessuna valutazione finora
- Lez 6 Fisiologia Umana 2020 2021 Urinario CarducciDocumento50 pagineLez 6 Fisiologia Umana 2020 2021 Urinario CarducciCaterina NuzzoNessuna valutazione finora
- Apparato UrinarioDocumento3 pagineApparato UrinariodanielaNessuna valutazione finora
- 9 10 Sistema UrogenitaleDocumento10 pagine9 10 Sistema UrogenitaleELENA ISABELA TAUTUNessuna valutazione finora
- Anatomia 2Documento90 pagineAnatomia 2Antonio Delle faveNessuna valutazione finora
- Interpretare Gli Esami Di LaboratorioDocumento177 pagineInterpretare Gli Esami Di LaboratoriogiancarloNessuna valutazione finora
- ReneDocumento83 pagineReneCaterina NuzzoNessuna valutazione finora
- Appunti Biochimica Del ReneDocumento57 pagineAppunti Biochimica Del ReneLuis CriscuoloNessuna valutazione finora
- Fegato e Vie BiliariDocumento14 pagineFegato e Vie BiliariDrHouse86Nessuna valutazione finora
- Apparato UrinarioDocumento11 pagineApparato UrinarioDani Lo MagroNessuna valutazione finora
- Nefrologia PDFDocumento182 pagineNefrologia PDFmaxNessuna valutazione finora
- Apparato EscretoreDocumento3 pagineApparato EscretoreGiacomo VatteroniNessuna valutazione finora
- APPARATO URINARIO (Salvato Automaticamente)Documento7 pagineAPPARATO URINARIO (Salvato Automaticamente)ENRICANessuna valutazione finora
- TRASPORTO TUBULAREDocumento4 pagineTRASPORTO TUBULAREHghh NgthhNessuna valutazione finora
- RIASSUNTO RENI (Cap 19-20)Documento4 pagineRIASSUNTO RENI (Cap 19-20)ANNALISA ZINNINessuna valutazione finora
- UrologiaDocumento51 pagineUrologiar2kmtyrdqdNessuna valutazione finora
- RENEDocumento85 pagineRENEGianluca LibianiNessuna valutazione finora
- Il Rene e La Struttura Del NefroneDocumento14 pagineIl Rene e La Struttura Del NefroneTONY TRUMPETNessuna valutazione finora
- Senza Nome 2 (Documento Riparato) .OdtDocumento6 pagineSenza Nome 2 (Documento Riparato) .OdtVanessaNessuna valutazione finora
- Rene BiologiaDocumento14 pagineRene BiologiaGiuseppe TomarchioNessuna valutazione finora
- Anatomia e Fisiologia Del Rene Prof - PapettiDocumento4 pagineAnatomia e Fisiologia Del Rene Prof - PapettiMattia NardoziNessuna valutazione finora
- Apparato UrinarioDocumento5 pagineApparato UrinarioPIPPORUBBATONessuna valutazione finora
- FisiologiaDocumento308 pagineFisiologiajessymsbNessuna valutazione finora
- Anatomia Apparato Urinario1Documento4 pagineAnatomia Apparato Urinario1FedericaRomanoNessuna valutazione finora
- Il Rene e La Struttura Del NefroneDocumento9 pagineIl Rene e La Struttura Del NefroneAntoniettaNessuna valutazione finora
- Dispensa Nefrologia PDFDocumento163 pagineDispensa Nefrologia PDFChiara VillanoNessuna valutazione finora
- Anatomia e Fisiologia Del ReneDocumento11 pagineAnatomia e Fisiologia Del ReneLorenzo FasanellaNessuna valutazione finora
- Fegato PancreasDocumento67 pagineFegato PancreasSergio100% (1)
- Anatomia e Fisiologia Del Rene Favaro SDocumento6 pagineAnatomia e Fisiologia Del Rene Favaro SMirelaNessuna valutazione finora
- Fisiologia Animal Parte 3Documento33 pagineFisiologia Animal Parte 3Vanda SheilaNessuna valutazione finora
- 2 - Urine e FeciDocumento31 pagine2 - Urine e Fecifrancesca.mazzilli63Nessuna valutazione finora
- UrologiaDocumento20 pagineUrologiaCarla OlivieroNessuna valutazione finora
- Du Dispense 2014Documento89 pagineDu Dispense 2014Cri EminaNessuna valutazione finora
- SCIENZEDocumento4 pagineSCIENZEandrea.mandolesiNessuna valutazione finora
- FEGATODocumento2 pagineFEGATOGiulia TestinoNessuna valutazione finora
- 38 Anatomia 18-11Documento9 pagine38 Anatomia 18-11Davide MarellaNessuna valutazione finora
- Apparato Digerente 2Documento15 pagineApparato Digerente 2melissaNessuna valutazione finora
- Anatomia II Lezione 27-05-2022Documento38 pagineAnatomia II Lezione 27-05-2022Asia GrassadoniaNessuna valutazione finora
- Sistema CirculatorioDocumento8 pagineSistema CirculatorioJusta MemeNessuna valutazione finora
- Il Sistema VascolareDocumento21 pagineIl Sistema VascolareNicolò Gino CoratoNessuna valutazione finora
- Appunti Apparato CardiocircolatorioDocumento11 pagineAppunti Apparato Cardiocircolatoriovirginia carparelliNessuna valutazione finora
- L'Apparato EscretoreDocumento2 pagineL'Apparato EscretoreVanny CorteseNessuna valutazione finora
- FARMACOLOGIA - Escrezione, Clearance e Interazioni Tra FarmaciDocumento12 pagineFARMACOLOGIA - Escrezione, Clearance e Interazioni Tra FarmaciChiaraNessuna valutazione finora
- Sistema EscretoreDocumento3 pagineSistema EscretoreDavideNessuna valutazione finora
- Dispensa Di Drenaggio Linfatico Manuale Per MCBDocumento12 pagineDispensa Di Drenaggio Linfatico Manuale Per MCBPaoloNessuna valutazione finora
- Apparato EscretoreDocumento5 pagineApparato EscretoreMilla KissNessuna valutazione finora
- Dispensa Anatomia CompletaDocumento19 pagineDispensa Anatomia CompletaVeronica ViolaNessuna valutazione finora