Potrebbero piacerti anche
- Lezione 06-FisiologiaDocumento3 pagineLezione 06-FisiologiaSarah ScuderiNessuna valutazione finora
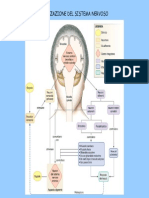
- Potenziale D'azioneDocumento8 paginePotenziale D'azioneMicheleNessuna valutazione finora
- Citologia 14 - Potenziale Di MembranaDocumento5 pagineCitologia 14 - Potenziale Di MembranaFrancesco TarulloNessuna valutazione finora
- Lezione 04-FisiologiaDocumento5 pagineLezione 04-FisiologiaSarah ScuderiNessuna valutazione finora
- Appunti Fisiologia IDocumento27 pagineAppunti Fisiologia Ikekko88Nessuna valutazione finora
- Canali Ionici, Fibre NervoseDocumento22 pagineCanali Ionici, Fibre Nervoseapi-3704348100% (1)
- Docsity Domande Triennale Fisiologia UmanaDocumento33 pagineDocsity Domande Triennale Fisiologia Umanadestinyema78Nessuna valutazione finora
- Potenziale D'azioneDocumento30 paginePotenziale D'azioneRiccardo Mason TroiaNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- 05 - I Canali Ionici - Parte Prima - 2015Documento10 pagine05 - I Canali Ionici - Parte Prima - 2015Nicky KellyNessuna valutazione finora
- Potenziale AzioneDocumento30 paginePotenziale AzionePatrizio SpadoniNessuna valutazione finora
- Fisiologia Lezione 4Documento6 pagineFisiologia Lezione 4The last chimeraNessuna valutazione finora
- Fisiologia Dei Sistemi MotoriDocumento17 pagineFisiologia Dei Sistemi Motorisara.zandonai99Nessuna valutazione finora
- Potenziali D'azioneDocumento5 paginePotenziali D'azioneHghh NgthhNessuna valutazione finora
- La Pompa Sodio-PotassioDocumento19 pagineLa Pompa Sodio-PotassioDavide ValmorbidaNessuna valutazione finora
- Logiche DinamicheDocumento3 pagineLogiche DinamicheMatteoBalzaniNessuna valutazione finora
- Lezione 6-MusumeciDocumento17 pagineLezione 6-MusumeciLorenzo CampanellaNessuna valutazione finora
- Fisiologia II-7 - Il Sistema Di ConduzioneDocumento14 pagineFisiologia II-7 - Il Sistema Di ConduzioneShaun Ivan MuzicNessuna valutazione finora
- FISIOLOGIAsbobineDocumento164 pagineFISIOLOGIAsbobinemarco.ricco.mr00Nessuna valutazione finora
- Lezione 2-MusumeciDocumento22 pagineLezione 2-MusumeciLorenzo CampanellaNessuna valutazione finora
- Conduzione Nervosa (Riassunto)Documento10 pagineConduzione Nervosa (Riassunto)Aaron RomanoNessuna valutazione finora
- 5 Potenziali ElettrotoniciDocumento8 pagine5 Potenziali ElettrotoniciClaudia OrlannoNessuna valutazione finora
- Conduzione NervosaDocumento1 paginaConduzione NervosaeurusneNessuna valutazione finora
- Il Potenziale D'azioneDocumento6 pagineIl Potenziale D'azioneLeandra EvolaNessuna valutazione finora
- Fisiologia Cellulare Potenziale AzioneDocumento31 pagineFisiologia Cellulare Potenziale Azionesimo_pNessuna valutazione finora
- 04 - Trasporto Di MembranaDocumento30 pagine04 - Trasporto Di MembranaÁurea RomãoNessuna valutazione finora
- Elettronica LCDocumento117 pagineElettronica LCIngegnere Pazzo100% (3)
- 02 Potenziale D'azioneDocumento13 pagine02 Potenziale D'azioneaxelshippoNessuna valutazione finora
- Fisiologia Parte 4.pdaDocumento14 pagineFisiologia Parte 4.pdaapi-3774792Nessuna valutazione finora
- Conduzione NervosaDocumento10 pagineConduzione NervosaMiriam LongobardiNessuna valutazione finora
- Anche Ad Altre ComponentiDocumento4 pagineAnche Ad Altre Componentifrancesco biancoNessuna valutazione finora
- NEUROBIOLOGIA La SapienzaDocumento30 pagineNEUROBIOLOGIA La Sapienzachiara cimeiNessuna valutazione finora
- 01-La Genesi Dell'impulso Nervoso PDFDocumento8 pagine01-La Genesi Dell'impulso Nervoso PDFDiego Flores EgüezNessuna valutazione finora
- Canali IoniciDocumento3 pagineCanali IoniciFilomena LongobardiNessuna valutazione finora
- Canali Ionici, Enzimi, Molecole CarrierDocumento114 pagineCanali Ionici, Enzimi, Molecole Carrierapi-26155364Nessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Segnali BioelettriciDocumento12 pagineSegnali BioelettriciFrancesco BorealeNessuna valutazione finora
- 4 - 07-03 Amp ErrorDocumento9 pagine4 - 07-03 Amp Errorortos11Nessuna valutazione finora
- Riassuntidi BiofisicaDocumento14 pagineRiassuntidi BiofisicaFlavia IovinoNessuna valutazione finora
- Processo Di Assorbimento (14marzo)Documento6 pagineProcesso Di Assorbimento (14marzo)Valerio CurcioNessuna valutazione finora
- 3 Equazione Di GoldmanDocumento2 pagine3 Equazione Di GoldmanClaudia OrlannoNessuna valutazione finora
- Nuova Parte 7 OndedurtoDocumento90 pagineNuova Parte 7 OndedurtobetriceNessuna valutazione finora
- Fisiologia - Riassunti Kandel "Principi Di Neuroscienze" Cap. 6-15Documento32 pagineFisiologia - Riassunti Kandel "Principi Di Neuroscienze" Cap. 6-15Matteo Paolucci100% (15)
- 3 - Trasporto AttivoDocumento9 pagine3 - Trasporto Attivoapi-26155364Nessuna valutazione finora
- Fisiologia 2Documento9 pagineFisiologia 2api-3774792Nessuna valutazione finora
- Trasporti Attraverso La MembranaDocumento36 pagineTrasporti Attraverso La MembranaSaraNessuna valutazione finora
- Os Cilla TBFDocumento13 pagineOs Cilla TBFmegadaveNessuna valutazione finora
- OperazionaliDocumento12 pagineOperazionaliMarco FunaroNessuna valutazione finora
- Legge Di GHK, Comparazione Membrana - Condensatore, Reobase e Cronassia, Canali, Canali Pre Il SodioDocumento33 pagineLegge Di GHK, Comparazione Membrana - Condensatore, Reobase e Cronassia, Canali, Canali Pre Il Sodioapi-3704348100% (2)
- Fisiologia Lezione 3Documento4 pagineFisiologia Lezione 3The last chimeraNessuna valutazione finora
- Lezione 9Documento11 pagineLezione 9Lorenzo CampanellaNessuna valutazione finora
- Rapporto Onde StazionarieDocumento6 pagineRapporto Onde StazionarieCristiano BruschiniNessuna valutazione finora
- Geologia Applicata 24 Novembre BisDocumento8 pagineGeologia Applicata 24 Novembre BisandreamercuriNessuna valutazione finora
- Quaderno - Elettronica Nicola Auricchio PDFDocumento292 pagineQuaderno - Elettronica Nicola Auricchio PDFYuriAmbrosio100% (1)
- Lezione Reattori ChimiciDocumento4 pagineLezione Reattori ChimiciAngelo Ngiò Francesco IovinoNessuna valutazione finora
- Lezione 08-FisiologiaDocumento5 pagineLezione 08-FisiologiaSarah ScuderiNessuna valutazione finora
- Lezione 10-FisiologiaDocumento5 pagineLezione 10-FisiologiaSarah ScuderiNessuna valutazione finora
- Lezione 09-FisiologiaDocumento3 pagineLezione 09-FisiologiaSarah ScuderiNessuna valutazione finora
- Potenziale Di MembranaDocumento4 paginePotenziale Di MembranaSimona CafàNessuna valutazione finora
- Lezione 01-FisiologiaDocumento1 paginaLezione 01-FisiologiaSarah ScuderiNessuna valutazione finora
- Bruniana & Campanelliana Vol. 9, No. 2, 2003 PDFDocumento270 pagineBruniana & Campanelliana Vol. 9, No. 2, 2003 PDFVetusta MaiestasNessuna valutazione finora
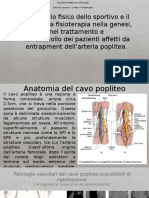
- Intrappolamento Dell'arteria Poplitea.Documento14 pagineIntrappolamento Dell'arteria Poplitea.AndreaGiustiNessuna valutazione finora
- Guida Insegnante VolentieriA2Documento49 pagineGuida Insegnante VolentieriA2Shreya ManojNessuna valutazione finora
- Il Contratto Di Noleggio e La Clausola CancelloDocumento2 pagineIl Contratto Di Noleggio e La Clausola CancelloAnna Paola SannaNessuna valutazione finora
- Da Muggia A TriesteDocumento47 pagineDa Muggia A TriesteRenzoMaggioreNessuna valutazione finora
- Salvatore Abbruzzese - Sociologia Delle ReligioniDocumento63 pagineSalvatore Abbruzzese - Sociologia Delle ReligioniDaniel MoranNessuna valutazione finora
- Edilizia in Legno-Linee Guida Toscana-Parte 1 Di 2Documento179 pagineEdilizia in Legno-Linee Guida Toscana-Parte 1 Di 2Mario Grimaudo100% (1)