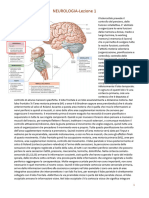
Potrebbero piacerti anche
- Occhiali a fori stenopeici: Come funzionano, come si usano, quali scegliereDa EverandOcchiali a fori stenopeici: Come funzionano, come si usano, quali scegliereNessuna valutazione finora
- Domandeleggio CORRETTEDocumento9 pagineDomandeleggio CORRETTEfrancescaNessuna valutazione finora
- NEUROPSICOLOGIA: le basi della materia (psicologia per tutti)Da EverandNEUROPSICOLOGIA: le basi della materia (psicologia per tutti)Valutazione: 3.5 su 5 stelle3.5/5 (2)
- Tutti UnitiDocumento120 pagineTutti UnitiERIKA BERNOCCHINessuna valutazione finora
- I bastoncelli retinici come cellule altamente specializzate nella fotorecezione : individuazione e caratterizzazione del metabolismo del cADPRDa EverandI bastoncelli retinici come cellule altamente specializzate nella fotorecezione : individuazione e caratterizzazione del metabolismo del cADPRNessuna valutazione finora
- Cariulo Sintonizzazione IntenzionaleDocumento14 pagineCariulo Sintonizzazione IntenzionaleFabrizio CastiNessuna valutazione finora
- Fondamenti Di Neuroscienze - Capitolo01Documento8 pagineFondamenti Di Neuroscienze - Capitolo01Elisa LR100% (1)
- Diagnosi e Correzione delle disfunzioni vertebrali con il percussore neuromuscolareDa EverandDiagnosi e Correzione delle disfunzioni vertebrali con il percussore neuromuscolareNessuna valutazione finora
- Giacomo Rizzolatti - Neuroni Specchio (Da Le Scienze N 460)Documento7 pagineGiacomo Rizzolatti - Neuroni Specchio (Da Le Scienze N 460)edosuper1100% (1)
- Giacomo Rizzolati - C'e Uno Specchio Nel Cervello - L'EspressoDocumento4 pagineGiacomo Rizzolati - C'e Uno Specchio Nel Cervello - L'EspressoVittoria Di BisceglieNessuna valutazione finora
- Relazione Del LibroDocumento8 pagineRelazione Del LibroPietro BottinoNessuna valutazione finora
- Riassunto Neuroni SpecchioDocumento18 pagineRiassunto Neuroni Specchiodaniele ferrariNessuna valutazione finora
- Lo Sguardo Degli AltriDocumento43 pagineLo Sguardo Degli AltriVincenzo CipollettaNessuna valutazione finora
- 1 FondamentiAFDocumento34 pagine1 FondamentiAFAxNessuna valutazione finora
- Guardare Gli Altri, Capire Gli Altri: 5.1 Il Movimento Non Va Pensato Una Concezione Antichissima Ha TenuDocumento5 pagineGuardare Gli Altri, Capire Gli Altri: 5.1 Il Movimento Non Va Pensato Una Concezione Antichissima Ha TenuNazariy DublianskyyNessuna valutazione finora
- Fisiologia VestibolareDocumento14 pagineFisiologia VestibolareAlinaNessuna valutazione finora
- Neuroni SpecchioDocumento3 pagineNeuroni Specchioangurp2575Nessuna valutazione finora
- 13 TELENCEFALO Sistema LimbicoDocumento2 pagine13 TELENCEFALO Sistema LimbicoRoberta PirasNessuna valutazione finora
- Neuroni SpecchioDocumento32 pagineNeuroni SpecchioPasqualeNessuna valutazione finora
- 6 Riflessi SpinaliDocumento38 pagine6 Riflessi SpinaliGiuseppe FoggiaNessuna valutazione finora
- PW 3Documento2 paginePW 3GiorgioNessuna valutazione finora
- Fondamenti Neuroscienze ComportamentoDocumento19 pagineFondamenti Neuroscienze ComportamentoRossanaNessuna valutazione finora
- Psicobiologia-1 371Documento117 paginePsicobiologia-1 371davideptraderNessuna valutazione finora
- Lezione 1Documento29 pagineLezione 1ylenia.alexandra.salaniNessuna valutazione finora
- 11-Sistema Nervoso 2017Documento91 pagine11-Sistema Nervoso 2017Murilo AlbuquerqueNessuna valutazione finora
- Psicofisiologia Delle Attivita' Mentali: Andrea PiarulliDocumento45 paginePsicofisiologia Delle Attivita' Mentali: Andrea PiarullimkuntzNessuna valutazione finora
- Fogassi Monza 6 Aprile 2020 Seconda ParteDocumento4 pagineFogassi Monza 6 Aprile 2020 Seconda ParteAnalía Figueroa CastelgrandeNessuna valutazione finora
- Lezione 15: Il Tessuto Nervoso IDocumento15 pagineLezione 15: Il Tessuto Nervoso IEmiliaNessuna valutazione finora
- Espressione CorporeaDocumento184 pagineEspressione CorporeaBenedetto RutiloNessuna valutazione finora
- I Neuroni A Specchio RIASSUNTODocumento11 pagineI Neuroni A Specchio RIASSUNTOGreta AllieriNessuna valutazione finora
- 7 - Il Sistema Nervoso 3Documento10 pagine7 - Il Sistema Nervoso 3Giulia BrancadoroNessuna valutazione finora
- Riassunti Di Anatomia 2 - Sistema Nervoso (Con SNV)Documento32 pagineRiassunti Di Anatomia 2 - Sistema Nervoso (Con SNV)giorgiocaresanaNessuna valutazione finora
- 2011 Funzioni EsecutiveDocumento87 pagine2011 Funzioni EsecutiveGabriele Sonnyboy OlivaNessuna valutazione finora
- Neuroscienze Cognitive ZanichelliDocumento275 pagineNeuroscienze Cognitive ZanichelliMaria DonatielloNessuna valutazione finora
- Principi NeuroscienzeDocumento12 paginePrincipi NeuroscienzegwalidorNessuna valutazione finora
- Intenzionalita' e Anosognosia PDFDocumento113 pagineIntenzionalita' e Anosognosia PDFNatalia MenottiNessuna valutazione finora
- Neuroni SpecchioDocumento2 pagineNeuroni SpecchioSimone AmericiNessuna valutazione finora
- Fondamenti Di Neuroscienze - Capitolo03Documento10 pagineFondamenti Di Neuroscienze - Capitolo03Elisa LR0% (1)
- Psicologia Dell AttenzioneDocumento101 paginePsicologia Dell AttenzioneMarco MenguzzatoNessuna valutazione finora
- Riassunto Libro Psicologia Generale Dal Cervello Alla Mente PDFDocumento19 pagineRiassunto Libro Psicologia Generale Dal Cervello Alla Mente PDFAlessandra Gulotta100% (1)
- Fondamenti Anatomo-Fisiologici Dei Processi PsichiciDocumento169 pagineFondamenti Anatomo-Fisiologici Dei Processi Psichicibeci8122003Nessuna valutazione finora
- Neurologia-11429Documento159 pagineNeurologia-11429Evelyn ReinosoNessuna valutazione finora
- La Simulazione Incarnata I Neuroni SpecchioDocumento9 pagineLa Simulazione Incarnata I Neuroni Specchion6m57psd89Nessuna valutazione finora
- Propriospinali Commissura: Midollo SpinaleDocumento1 paginaPropriospinali Commissura: Midollo SpinaleP.Nessuna valutazione finora
- Appunti Psicologia Dello SportDocumento22 pagineAppunti Psicologia Dello SportdanidragosportNessuna valutazione finora
- Lezione EmpatiaDocumento6 pagineLezione EmpatiaSara BuonadosaNessuna valutazione finora
- Sistema SensorimotorioDocumento6 pagineSistema SensorimotorioMiriam LongobardiNessuna valutazione finora
- Fondamenti Di Neuroscienze - Capitolo02Documento8 pagineFondamenti Di Neuroscienze - Capitolo02Elisa LRNessuna valutazione finora
- 9 Corteccia MotoriaDocumento39 pagine9 Corteccia MotoriaMichele MarengoNessuna valutazione finora
- Presentazione Sistema NervosoDocumento60 paginePresentazione Sistema NervosoMattNessuna valutazione finora
- NeuropsicologiaDocumento54 pagineNeuropsicologiaNoura AllaouiNessuna valutazione finora
- Reti NeuraliDocumento17 pagineReti Neuralialberto_giudici6672100% (1)
- Sistema Nervoso Centrale e PerifericoDocumento11 pagineSistema Nervoso Centrale e PerifericoELENA ISABELA TAUTUNessuna valutazione finora
- Neuroanatomia (PG)Documento82 pagineNeuroanatomia (PG)Sio88100% (2)
- 1 A Introd ABADocumento14 pagine1 A Introd ABAAntonioNessuna valutazione finora
- Sistema Nervoso2Documento41 pagineSistema Nervoso2Agostino InzerilloNessuna valutazione finora
- Neurofisiologia FilesDocumento13 pagineNeurofisiologia FilesAlessandroE.E.FrancoNessuna valutazione finora
- Reti Neurali - ScalettaDocumento1 paginaReti Neurali - Scalettaalberto_giudici6672Nessuna valutazione finora
- NeuropsicologiaDocumento126 pagineNeuropsicologiaBeatriceNessuna valutazione finora
- So Quel Che FaiDocumento3 pagineSo Quel Che FaiSonia Pisani0% (1)
- Sostituzione UovaDocumento1 paginaSostituzione UovaSonia PisaniNessuna valutazione finora
- La Terapia Amniotica Con I Pazienti Grav PDFDocumento3 pagineLa Terapia Amniotica Con I Pazienti Grav PDFSonia PisaniNessuna valutazione finora
- Schema Per L'analisi GrammaticaleDocumento4 pagineSchema Per L'analisi GrammaticaleIticescu Diana63% (8)
- Ciambella Allo ZenzeroDocumento1 paginaCiambella Allo ZenzeroSonia PisaniNessuna valutazione finora
- Cheesecake Alle FragoleDocumento2 pagineCheesecake Alle FragoleSonia PisaniNessuna valutazione finora
- I Diritti Dei Cittadini Con DisabilitàDocumento46 pagineI Diritti Dei Cittadini Con DisabilitàSonia PisaniNessuna valutazione finora
- Focaccia Al PomodoroDocumento1 paginaFocaccia Al PomodoroSonia PisaniNessuna valutazione finora
- Seitan Limone e AlloroDocumento1 paginaSeitan Limone e AlloroSonia PisaniNessuna valutazione finora
- Alcune RicetteDocumento64 pagineAlcune Ricettemaracaibo1984Nessuna valutazione finora
- Spagiria e AchimiaDocumento62 pagineSpagiria e AchimiaIvana MarquezNessuna valutazione finora
- Brasato Di SeitanDocumento1 paginaBrasato Di SeitanSonia PisaniNessuna valutazione finora
- Psicologia Sociale - L'aggressivitàDocumento10 paginePsicologia Sociale - L'aggressivitàMatteoRagneddaNessuna valutazione finora
- Sine Requie EditabileDocumento1 paginaSine Requie EditabileMitic93Nessuna valutazione finora
- Pragmatica Della Comunicazione Umana Processi e Dinamiche Di GruppoDocumento35 paginePragmatica Della Comunicazione Umana Processi e Dinamiche Di GruppoChiara BreciNessuna valutazione finora
- Appunti Topografia-Idoneità-4°Documento43 pagineAppunti Topografia-Idoneità-4°salvoNessuna valutazione finora
- Il Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoDa EverandIl Grande Dizionario della MetaMedicina: Guarire interpretando i messaggi del corpoNessuna valutazione finora
- Anatomia della Guarigione: I sette principi della Nuova Medicina IntegrataDa EverandAnatomia della Guarigione: I sette principi della Nuova Medicina IntegrataValutazione: 5 su 5 stelle5/5 (2)
- Anatomia della Coscienza Quantica: La fisica dell'auto-guarigioneDa EverandAnatomia della Coscienza Quantica: La fisica dell'auto-guarigioneValutazione: 4.5 su 5 stelle4.5/5 (3)
- Psicologia Oscura: Scopri come analizzare le persone e padroneggiare la manipolazione umana utilizzando i segreti del linguaggio del corpo, la PNL sottile, il controllo mentale, la persuasione subliminale, l'ipnosi e le tecniche di lettura veloce.Da EverandPsicologia Oscura: Scopri come analizzare le persone e padroneggiare la manipolazione umana utilizzando i segreti del linguaggio del corpo, la PNL sottile, il controllo mentale, la persuasione subliminale, l'ipnosi e le tecniche di lettura veloce.Valutazione: 5 su 5 stelle5/5 (6)
- Dall'amore cieco all'amore consapevole. Relazione tra la pedagogia e le costellazioni di Bert HellingerDa EverandDall'amore cieco all'amore consapevole. Relazione tra la pedagogia e le costellazioni di Bert HellingerNessuna valutazione finora
- Ricomincia dalla pancia: Scopri il potere anti-età del microbioma e ritrova la tua salute partendo da dentroDa EverandRicomincia dalla pancia: Scopri il potere anti-età del microbioma e ritrova la tua salute partendo da dentroIsabella PolliNessuna valutazione finora
- Le Tattiche del Cambiamento - Manuale di Psicoterapia StrategicaDa EverandLe Tattiche del Cambiamento - Manuale di Psicoterapia StrategicaValutazione: 5 su 5 stelle5/5 (1)
- La medicina di santa Ildegarda: Per tutti i giorni e per tutta la famiglia.Da EverandLa medicina di santa Ildegarda: Per tutti i giorni e per tutta la famiglia.Valutazione: 5 su 5 stelle5/5 (2)
- Gonfiore Addominale: Guide e consigli per contrastare la pancia gonfiaDa EverandGonfiore Addominale: Guide e consigli per contrastare la pancia gonfiaNessuna valutazione finora
- Piena Coscienza : Lezioni da leggere prima di dormireDa EverandPiena Coscienza : Lezioni da leggere prima di dormireNessuna valutazione finora
- La Mentalità Di Successo Dei Grandi Leader: Non è il denaroDa EverandLa Mentalità Di Successo Dei Grandi Leader: Non è il denaroNessuna valutazione finora
- Il Piano Dietetico del Dr. Nowzaradan: La Bilancia non Mente, la Gente Mente! L'unica Dieta da 1200 kcal del Dr NOW per Perdere Peso Velocemente. Piano Dietetico di 30 GiorniDa EverandIl Piano Dietetico del Dr. Nowzaradan: La Bilancia non Mente, la Gente Mente! L'unica Dieta da 1200 kcal del Dr NOW per Perdere Peso Velocemente. Piano Dietetico di 30 GiorniNessuna valutazione finora
- Sindrome di Asperger e Counselling - Importanza dell’Intervento di Supporto nel Processo di Adattamento all’Interno dell’Ambiente LavorativoDa EverandSindrome di Asperger e Counselling - Importanza dell’Intervento di Supporto nel Processo di Adattamento all’Interno dell’Ambiente LavorativoNessuna valutazione finora
- Prevenire le Malattie: Strategie Rivoluzionarie per un Domani più Sano: Potenziare il Tuo Percorso verso il Benessere attraverso la Scienza e l'InnovazioneDa EverandPrevenire le Malattie: Strategie Rivoluzionarie per un Domani più Sano: Potenziare il Tuo Percorso verso il Benessere attraverso la Scienza e l'InnovazioneNessuna valutazione finora
- Identità Rara. La lotta dei malati rari per il riconoscimento socialeDa EverandIdentità Rara. La lotta dei malati rari per il riconoscimento socialeNessuna valutazione finora
- I due volti dell’amore: Come far funzionare l'amore nei rapporti affettiviDa EverandI due volti dell’amore: Come far funzionare l'amore nei rapporti affettiviNessuna valutazione finora
- Cambia la tua vita con la Legge di Attrazione, ma prima sblocca le credenze limitanti che ti hanno sempre impedito di farla funzionare!: Ma prima sblocca le Credenze limitanti che ti hanno sempre impedito di farla funzionare!Da EverandCambia la tua vita con la Legge di Attrazione, ma prima sblocca le credenze limitanti che ti hanno sempre impedito di farla funzionare!: Ma prima sblocca le Credenze limitanti che ti hanno sempre impedito di farla funzionare!Nessuna valutazione finora
- L’altra faccia della luna: Emozioni e PsicoanalisiDa EverandL’altra faccia della luna: Emozioni e PsicoanalisiNessuna valutazione finora
- Il miracolo della medicina rigenerativa: Come invertire il processo d’invecchiamento in modo naturaleDa EverandIl miracolo della medicina rigenerativa: Come invertire il processo d’invecchiamento in modo naturaleNessuna valutazione finora
- La vera agopuntura cinese - Dottrina, Diagnosi, TerapiaDa EverandLa vera agopuntura cinese - Dottrina, Diagnosi, TerapiaNessuna valutazione finora