Potrebbero piacerti anche
- Frutti Antichi - Antiche Varietà LecchesiDocumento145 pagineFrutti Antichi - Antiche Varietà Lecchesicontadino_impazzitoNessuna valutazione finora
- Microbiologia Degli Alimenti (Testo)Documento80 pagineMicrobiologia Degli Alimenti (Testo)Paolo GeiNessuna valutazione finora
- EM Libro HigaDocumento11 pagineEM Libro Higaascionefrancesco85100% (1)
- Bioindicatori e BiomarkersDocumento42 pagineBioindicatori e Biomarkersdadi1964Nessuna valutazione finora
- Forme Di Allevamento - Vaso Catalano - DiapositiveDocumento21 pagineForme Di Allevamento - Vaso Catalano - Diapositivecontadino_impazzitoNessuna valutazione finora
- BiotecnologiaDocumento18 pagineBiotecnologiamelissa lopezNessuna valutazione finora
- Lotta Biologica in AgricolturaDocumento134 pagineLotta Biologica in Agricolturapedro41Nessuna valutazione finora
- Report Pesticidi Salute 2022Documento25 pagineReport Pesticidi Salute 2022Francesco SchioppaNessuna valutazione finora
- Agricoltura Sostenibile (Ecocompatibile) Le Filiere AgroalimentariDocumento64 pagineAgricoltura Sostenibile (Ecocompatibile) Le Filiere AgroalimentariDomenico Bevilacqua50% (2)
- Biologia VegetaleDocumento21 pagineBiologia VegetaleUomo RagnoNessuna valutazione finora
- Chimica TossicologicaDocumento72 pagineChimica TossicologicapaolachitNessuna valutazione finora
- Gestione Sostenibile Delle Erbe InfestantiDocumento34 pagineGestione Sostenibile Delle Erbe InfestantipiripicchioNessuna valutazione finora
- Articolo Biologico Cosmos KosmeticaDocumento4 pagineArticolo Biologico Cosmos Kosmeticaelissa.astarteNessuna valutazione finora
- L'agricoltura Biologica Stefano CanaliDocumento9 pagineL'agricoltura Biologica Stefano CanaliSamNessuna valutazione finora
- Piante Medicinali in Fitoterapia PDFDocumento6 paginePiante Medicinali in Fitoterapia PDFrodrigue yanNessuna valutazione finora
- Balcone FioritoDocumento4 pagineBalcone FioritoD3st1nyNessuna valutazione finora
- 1.patogenicita' e VirulenzaDocumento51 pagine1.patogenicita' e VirulenzaFrancesco SchioppaNessuna valutazione finora
- Aap Bioindicatori AmbientaliDocumento3 pagineAap Bioindicatori AmbientaliPier Luigi CavalchiniNessuna valutazione finora
- Piante Medicinali in FitoterapiaDocumento6 paginePiante Medicinali in FitoterapiaSergioNessuna valutazione finora
- Appunti Microbiologia Degli AlimentiDocumento129 pagineAppunti Microbiologia Degli AlimentiAntonio ScarpatoNessuna valutazione finora
- Il monitoraggio ambientale: Un efficace strumento di controlloDa EverandIl monitoraggio ambientale: Un efficace strumento di controlloNessuna valutazione finora
- Attività Antiossidante, Antimutagena e Antigenotossica dei LattobacilliDa EverandAttività Antiossidante, Antimutagena e Antigenotossica dei LattobacilliNessuna valutazione finora
- Metodologie Sintetiche EcosostenibiliDocumento143 pagineMetodologie Sintetiche EcosostenibiliMaria Cristina PesolaNessuna valutazione finora
- L ' Erboristeria VeterinariaDocumento16 pagineL ' Erboristeria VeterinariapippoNessuna valutazione finora
- Bioetica e OgmDocumento3 pagineBioetica e OgmEnrico BoattinNessuna valutazione finora
- Metodi Di Produzione A Basso Impatto AmbientaleDocumento3 pagineMetodi Di Produzione A Basso Impatto AmbientaleSautè AleNessuna valutazione finora
- Appunti Microbiologia Generale e Enologica PDFDocumento87 pagineAppunti Microbiologia Generale e Enologica PDFMarco Lami100% (1)
- Antecedentes Del TemaDocumento4 pagineAntecedentes Del TemaCarla BarriosNessuna valutazione finora
- PRO E CONTRO BiotecnologieDocumento2 paginePRO E CONTRO BiotecnologieSonia Putrino75% (4)
- Manuale BiocompostDocumento48 pagineManuale BiocompostFrancis ParthenicumNessuna valutazione finora
- Le Biotecnologie - I FermentatoriDocumento10 pagineLe Biotecnologie - I FermentatoriTeresa CicconeNessuna valutazione finora
- Qualità Dei Prodotti AnimaliDocumento60 pagineQualità Dei Prodotti AnimaliCaelo0Nessuna valutazione finora
- Docsity Agricoltura Slow Riassunto SottileDocumento38 pagineDocsity Agricoltura Slow Riassunto SottilesaegvfaefNessuna valutazione finora
- LesimbiosimicorrizicheDocumento9 pagineLesimbiosimicorrizicheCaterina StrafalaciNessuna valutazione finora
- Coltivare Funghi Commestibili: Guida Pratica con 20 Capitoli DettagliatiDa EverandColtivare Funghi Commestibili: Guida Pratica con 20 Capitoli DettagliatiNessuna valutazione finora
- Spiegazione FanghiDocumento7 pagineSpiegazione FanghiDaniela RuggeriNessuna valutazione finora
- LindanoDocumento68 pagineLindanobamma lasalungaNessuna valutazione finora
- La Chimica VerdeDocumento15 pagineLa Chimica VerdeMaria Cristina PesolaNessuna valutazione finora
- Le BiotecnologieDocumento3 pagineLe Biotecnologiegrecoanastasia731Nessuna valutazione finora
- Estrazione Di Metaboliti Secondari Della Pianta Brunfelsia Chirikaspi Plowman TESIDocumento35 pagineEstrazione Di Metaboliti Secondari Della Pianta Brunfelsia Chirikaspi Plowman TESIJorge Eduardo Ponce ZeaNessuna valutazione finora
- Introduzione Alla MicrobiologiaDocumento42 pagineIntroduzione Alla Microbiologiamaria_ponticelliNessuna valutazione finora
- AA - VV. - Compendio Di Farmacologia PDFDocumento270 pagineAA - VV. - Compendio Di Farmacologia PDFcoloromaneNessuna valutazione finora
- 001introduzione MicrobiologiaDocumento50 pagine001introduzione MicrobiologiaAlessandra Patricia LopesNessuna valutazione finora
- 1c Avversità Biotiche e Possibilità Di Intervento, SostenibilitàDocumento110 pagine1c Avversità Biotiche e Possibilità Di Intervento, SostenibilitàCarlos GolbanoNessuna valutazione finora
- 10indicatori PDFDocumento10 pagine10indicatori PDFDiego Flores EgüezNessuna valutazione finora
- Elementi Base Di Cosmetologia Naturale Parte IIDocumento35 pagineElementi Base Di Cosmetologia Naturale Parte IIfrancy_feaNessuna valutazione finora
- Perspectivas de Control BiologicoDocumento44 paginePerspectivas de Control BiologicoIKER NERINessuna valutazione finora
- Determinator SpeciiDocumento68 pagineDeterminator SpeciiDoina PopaNessuna valutazione finora
- La GenéticaDocumento3 pagineLa GenéticaSaima Becerra HurtadoNessuna valutazione finora
- 1212 LaChimicadelRestauroDocumento20 pagine1212 LaChimicadelRestauroLegambiente Circolo NebrodiNessuna valutazione finora
- Il Problema Etico Delle BiotecnologieDocumento3 pagineIl Problema Etico Delle BiotecnologieMartina FestaNessuna valutazione finora
- ..-Area - Protetta-Ceirsa - LINEE GUIDA PER UN CORRETTO CONTROLLO DELLE INFESTAZIONI DA INSETTI E ANIMALI INDESIDERATI - Def - Senza - Pipis PDFDocumento58 pagine..-Area - Protetta-Ceirsa - LINEE GUIDA PER UN CORRETTO CONTROLLO DELLE INFESTAZIONI DA INSETTI E ANIMALI INDESIDERATI - Def - Senza - Pipis PDFstefania matera0% (1)
- Dispensa 2020Documento23 pagineDispensa 2020Claudio CelestiniNessuna valutazione finora
- Riassunto Sugli OgmDocumento115 pagineRiassunto Sugli OgmRiccardo RicciNessuna valutazione finora
- Il GlifosatoDocumento3 pagineIl GlifosatoFrancesco FalconiNessuna valutazione finora
- Trattato Sul GanodermaDocumento24 pagineTrattato Sul Ganodermahscappa8519Nessuna valutazione finora
- Ab31 Ord19Documento3 pagineAb31 Ord19caligiuri.dinaNessuna valutazione finora
- Ciano Batter IDocumento97 pagineCiano Batter IFernando DuranteNessuna valutazione finora
- Considerazioni Sulla Proposta Della Commissione Europea Di Un Nuovo Regolamento Sull'uso Sostenibile Dei Prodotti FitosanitariDocumento6 pagineConsiderazioni Sulla Proposta Della Commissione Europea Di Un Nuovo Regolamento Sull'uso Sostenibile Dei Prodotti FitosanitariNatalino GuisoNessuna valutazione finora
- Tesi Giovanni Tuccio Per PDCDocumento130 pagineTesi Giovanni Tuccio Per PDCjonaNessuna valutazione finora
- Aspetti Giuridici e Deontologici 2 OkDocumento33 pagineAspetti Giuridici e Deontologici 2 OkIvanNessuna valutazione finora
- Micotossine Negli AlimentiDocumento57 pagineMicotossine Negli AlimentiMarco MacciNessuna valutazione finora
- Fertilizzanti Biostimolanti - Prova XurianDocumento9 pagineFertilizzanti Biostimolanti - Prova Xuriancontadino_impazzitoNessuna valutazione finora
- Sperimentazione Pomodoro Da IndustriaDocumento13 pagineSperimentazione Pomodoro Da Industriacontadino_impazzitoNessuna valutazione finora
- Manuale Per La Formazione Dell'Insegnante - Educazione Alimentare e Del Gusto Nella Scuola PrimariaDocumento163 pagineManuale Per La Formazione Dell'Insegnante - Educazione Alimentare e Del Gusto Nella Scuola Primariacontadino_impazzitoNessuna valutazione finora
- Fertilizzanti Innovativi - Bio Aksxter - Relazione TecnicaDocumento8 pagineFertilizzanti Innovativi - Bio Aksxter - Relazione Tecnicacontadino_impazzitoNessuna valutazione finora
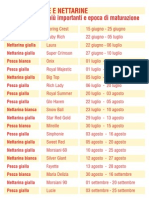
- Le Varietà Delle Pesche Nettarine - SchedaDocumento1 paginaLe Varietà Delle Pesche Nettarine - Schedacontadino_impazzitoNessuna valutazione finora
- Orticoltura - La Luna e L'Influenza Sulle ColtureDocumento7 pagineOrticoltura - La Luna e L'Influenza Sulle Colturecontadino_impazzitoNessuna valutazione finora
- Frutticoltura - Irrigazione in CerasicolturaDocumento4 pagineFrutticoltura - Irrigazione in Cerasicolturacontadino_impazzitoNessuna valutazione finora
- Leguminose Da Foraggio - DiapositiveDocumento91 pagineLeguminose Da Foraggio - Diapositivecontadino_impazzitoNessuna valutazione finora
- La Pacciamatura Nell'orto FamilareDocumento4 pagineLa Pacciamatura Nell'orto Familarecontadino_impazzitoNessuna valutazione finora
- Vantaggi Del SovescioDocumento6 pagineVantaggi Del Sovesciocontadino_impazzitoNessuna valutazione finora
- Fertilizzate Biostimolante - Xurian PDFDocumento2 pagineFertilizzate Biostimolante - Xurian PDFcontadino_impazzitoNessuna valutazione finora
- Il Pesco in Agricoltura BiologicaDocumento14 pagineIl Pesco in Agricoltura Biologicacontadino_impazzitoNessuna valutazione finora
- SORGO (Sorghum SPP.) - Scheda TecnicaDocumento2 pagineSORGO (Sorghum SPP.) - Scheda Tecnicacontadino_impazzitoNessuna valutazione finora
- Le Forme Di Allevamento ArboreeDocumento39 pagineLe Forme Di Allevamento Arboreecontadino_impazzitoNessuna valutazione finora
- Pesco, I Possibili Rimedi Ai Problemi Di RistoppioDocumento3 paginePesco, I Possibili Rimedi Ai Problemi Di Ristoppiocontadino_impazzitoNessuna valutazione finora
- Piante Usate in Italia in Medicina PopolareDocumento20 paginePiante Usate in Italia in Medicina Popolarecontadino_impazzitoNessuna valutazione finora
- Rotazioni in Agricoltura Biologica CRADocumento2 pagineRotazioni in Agricoltura Biologica CRAcontadino_impazzitoNessuna valutazione finora
- Strategie Di Controllo NematodiDocumento52 pagineStrategie Di Controllo Nematodicontadino_impazzitoNessuna valutazione finora
- Rotazioni Orticole in Agricoltura BiologicaDocumento20 pagineRotazioni Orticole in Agricoltura Biologicacontadino_impazzitoNessuna valutazione finora
- Brassicacee Biofumiganti e Salute Delle ColtureDocumento1 paginaBrassicacee Biofumiganti e Salute Delle Colturecontadino_impazzitoNessuna valutazione finora
- Sorgo e SiccitàDocumento3 pagineSorgo e Siccitàcontadino_impazzitoNessuna valutazione finora
- Sovesci Primaverili Ed EstiviDocumento12 pagineSovesci Primaverili Ed Estivicontadino_impazzitoNessuna valutazione finora
- Glicosidi CianogeneticiDocumento44 pagineGlicosidi Cianogeneticicontadino_impazzitoNessuna valutazione finora
- Ristoppio in FrutticolturaDocumento2 pagineRistoppio in Frutticolturacontadino_impazzito100% (1)
- Manuale Ulivo PDFDocumento145 pagineManuale Ulivo PDFcontadino_impazzitoNessuna valutazione finora
- Schede Tecniche Il Cipresso - Ideaverde - Vivai A PerugiaDocumento9 pagineSchede Tecniche Il Cipresso - Ideaverde - Vivai A PerugiaGinoNessuna valutazione finora
- ZIGOMICETI e DERMATOFITIDocumento10 pagineZIGOMICETI e DERMATOFITILiv TartaglioneNessuna valutazione finora
- Dispensa Corso MicologicoDocumento51 pagineDispensa Corso MicologicoWalter LentiniNessuna valutazione finora
- E: Ebook - Erbe - Clark PDFDocumento18 pagineE: Ebook - Erbe - Clark PDFElena MandaNessuna valutazione finora