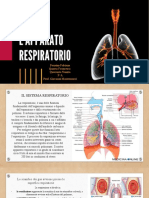
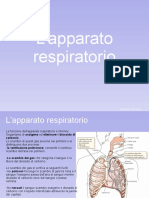
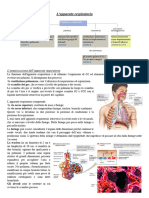
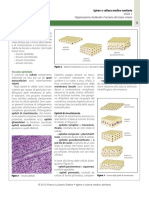
Potrebbero piacerti anche
- Glossario Istologia Ed EmbriologiaDocumento10 pagineGlossario Istologia Ed EmbriologiaVictoria CruzNessuna valutazione finora
- Sadava Nuovabiologiablu Sintesi Capc3 Plus PDFDocumento2 pagineSadava Nuovabiologiablu Sintesi Capc3 Plus PDFGabriele FasoloNessuna valutazione finora
- Guida Riconoscimento TessutiDocumento15 pagineGuida Riconoscimento Tessutimeneg90Nessuna valutazione finora
- Lez 5 Fisiologia Umana 2020 2021 Respiratorio CarducciDocumento85 pagineLez 5 Fisiologia Umana 2020 2021 Respiratorio CarducciCristian ChirNessuna valutazione finora
- Fisiologia Dell'apparato RespiratorioDocumento74 pagineFisiologia Dell'apparato RespiratorioShake PuzzaNessuna valutazione finora
- Apparato RespiratorioDocumento18 pagineApparato RespiratoriomelissaNessuna valutazione finora
- Scienze Apparato RespiratorioDocumento4 pagineScienze Apparato Respiratorioxgjp747zdxNessuna valutazione finora
- 17 - Apparato RespiratorioDocumento37 pagine17 - Apparato RespiratorioSara VerdoneNessuna valutazione finora
- 5 Il Sistema RespiratorioDocumento52 pagine5 Il Sistema Respiratorioas asNessuna valutazione finora
- Apparato RespiratorioDocumento13 pagineApparato RespiratorioStefano NicastroNessuna valutazione finora
- UntitledDocumento3 pagineUntitledRoberta ArmatoNessuna valutazione finora
- L'apparato RespiratorioDocumento31 pagineL'apparato RespiratorioElena Bandera100% (1)
- Apparato RespiratorioDocumento10 pagineApparato RespiratorioRim Taoufik El idrissiNessuna valutazione finora
- Il Sistema RespiratorioDocumento12 pagineIl Sistema RespiratorioFabiana FonsinoNessuna valutazione finora
- Biologia Capitolo C3Documento3 pagineBiologia Capitolo C3Denise NotaNessuna valutazione finora
- Apparato RespiratorioDocumento3 pagineApparato RespiratorioGiulia GiuffridaNessuna valutazione finora
- Apparato RespiratorioDocumento27 pagineApparato RespiratorioNicola RainoneNessuna valutazione finora
- Apparato RespiratorioDocumento2 pagineApparato RespiratorioGelsomino GrilloNessuna valutazione finora
- Seconda Annualità 1 Nucleo Tematico 1 Lezione-Gli Organi Dellapparato RespiratorioDocumento34 pagineSeconda Annualità 1 Nucleo Tematico 1 Lezione-Gli Organi Dellapparato Respiratoriopsnkw4d7wnNessuna valutazione finora
- Apparato RespiratorioDocumento4 pagineApparato RespiratoriouifdvhsidNessuna valutazione finora
- Apparato RespiratorioDocumento4 pagineApparato RespiratorioFrancesca FrizziNessuna valutazione finora
- Apparato RespiratorioDocumento9 pagineApparato Respiratoriochiara9rossoNessuna valutazione finora
- Fisiologia Respiratoria (Meccanica, Scambio e Trasporto Dei Gasi, Controllo Nervoso) PDFDocumento13 pagineFisiologia Respiratoria (Meccanica, Scambio e Trasporto Dei Gasi, Controllo Nervoso) PDFBobbyGustafson100% (1)
- Apparato RespiratorioDocumento5 pagineApparato RespiratorioCarlotta MazzoniNessuna valutazione finora
- Apparato Respiratorio PDFDocumento4 pagineApparato Respiratorio PDFnodlesNessuna valutazione finora
- Sistema RespiratorioDocumento4 pagineSistema RespiratorioIsser Mota bautistaNessuna valutazione finora
- Marieb Sintesi 62091 c9Documento2 pagineMarieb Sintesi 62091 c9aliarare22Nessuna valutazione finora
- Apparato RespiratorioDocumento13 pagineApparato RespiratorioRosa NoceraNessuna valutazione finora
- Sistema Respiratorio AniaDocumento3 pagineSistema Respiratorio AniaJavier GutierrezNessuna valutazione finora
- 14 Apparato RespiratorioDocumento10 pagine14 Apparato Respiratoriomariannaguadagno00Nessuna valutazione finora
- - apparato respiratorio e cardiovascolareDocumento29 pagine- apparato respiratorio e cardiovascolareColorado Express YTNessuna valutazione finora
- Fisiologia Apparato RespiratorioDocumento27 pagineFisiologia Apparato RespiratorioCristina Ivaldi50% (2)
- Malattie App. RespiratorioDocumento23 pagineMalattie App. RespiratorioMariateresa SantorsolaNessuna valutazione finora
- Apparato RespiratorioDocumento2 pagineApparato RespiratorioChiara CampisiNessuna valutazione finora
- ARDS Sindrome Da Distress Respiratorio AcutoDocumento20 pagineARDS Sindrome Da Distress Respiratorio AcutoPaolo FineNessuna valutazione finora
- Scienze Gaia FiorettoDocumento3 pagineScienze Gaia FiorettoGaia FiorettoNessuna valutazione finora
- Il Sistema RespiratorioDocumento15 pagineIl Sistema RespiratorioCaterina RizzoNessuna valutazione finora
- Scienze Motorie Francesco Montebello PDFDocumento5 pagineScienze Motorie Francesco Montebello PDFFrancesco MontebelloNessuna valutazione finora
- Apparato RespiratorioDocumento27 pagineApparato RespiratorioDile SimeNessuna valutazione finora
- Apparato RespiratorioDocumento3 pagineApparato RespiratorioCatalina Valeria MocanuNessuna valutazione finora
- Apparato RespiratorioDocumento20 pagineApparato RespiratorioFrancesco FabianiNessuna valutazione finora
- Mappa Concettuale Sull'Apparato RespiratorioDocumento1 paginaMappa Concettuale Sull'Apparato RespiratorioBOLFRA100% (1)
- Apparato RespiratorioDocumento6 pagineApparato RespiratorioPIPPORUBBATONessuna valutazione finora
- Sentido Del OlfatoDocumento13 pagineSentido Del OlfatoGustavo OrdoñezNessuna valutazione finora
- Anatomia 5Documento10 pagineAnatomia 5Roberto MekhailNessuna valutazione finora
- Fisiologia Dell'Apparato RespiratorioDocumento9 pagineFisiologia Dell'Apparato RespiratorioEleonora Francesco Consalvo DonzelliNessuna valutazione finora
- Apparato Respiratorio PDFDocumento18 pagineApparato Respiratorio PDFmaddalenaNessuna valutazione finora
- La Respirazione e L'apparato RespiratorioDocumento4 pagineLa Respirazione e L'apparato RespiratorioGiftNessuna valutazione finora
- Apparato RespiratorioDocumento10 pagineApparato RespiratorioDanilo SalonnaNessuna valutazione finora
- Apparato RespiratorioDocumento19 pagineApparato Respiratoriovaleria trujilloNessuna valutazione finora
- Sistema RespiratorioDocumento4 pagineSistema RespiratorioVioleta PérezNessuna valutazione finora
- Apparato RespiratorioDocumento1 paginaApparato RespiratorioMarta NataliniNessuna valutazione finora
- Apparato RespiratorioDocumento3 pagineApparato Respiratoriosara roccoNessuna valutazione finora
- Fisioterapia RespiratoriaDocumento56 pagineFisioterapia RespiratoriaFabrizio PaglierucciNessuna valutazione finora
- App. RespiratorioDocumento3 pagineApp. RespiratorioLaura Cornacchia RomeroNessuna valutazione finora
- RianimazioneDocumento33 pagineRianimazioneSimone SoccalNessuna valutazione finora
- RespirazioneDocumento18 pagineRespirazionemisesusNessuna valutazione finora
- Apparato RespiratorioDocumento4 pagineApparato RespiratorioMichele CasiniNessuna valutazione finora
- Pranayama, la scienza del respiro (translated)Da EverandPranayama, la scienza del respiro (translated)Valutazione: 4.5 su 5 stelle4.5/5 (2)
- Ginnastica respiratoria - Purificazione - Salute - Forza - Energia mediante l'esercizio fisico e psichico della respirazioneDa EverandGinnastica respiratoria - Purificazione - Salute - Forza - Energia mediante l'esercizio fisico e psichico della respirazioneValutazione: 4 su 5 stelle4/5 (1)
- Zanzare in Appenino - I culicidi di alta quota in provincia di ModenaDa EverandZanzare in Appenino - I culicidi di alta quota in provincia di ModenaNessuna valutazione finora
- Fisiologia RespiratoriaDocumento14 pagineFisiologia RespiratoriaJohnNessuna valutazione finora
- Elementi Di ECGDocumento28 pagineElementi Di ECGJohnNessuna valutazione finora
- AbcdeDocumento10 pagineAbcdeJohnNessuna valutazione finora
- Interpretazione Dell'Emogasanalisi: IRC (Italian Resuscitation Council)Documento17 pagineInterpretazione Dell'Emogasanalisi: IRC (Italian Resuscitation Council)JohnNessuna valutazione finora
- Epiteli SensorialiDocumento5 pagineEpiteli Sensorialiapi-3704348100% (1)
- Biologia 2Documento11 pagineBiologia 2Noemi ArzilliNessuna valutazione finora
- I TessutiDocumento19 pagineI TessutiGiorgioNessuna valutazione finora
- TESSUTIDocumento54 pagineTESSUTIercoleNessuna valutazione finora
- Patologia Forense Diapo 2Documento255 paginePatologia Forense Diapo 2MariaHoffman100% (1)
- Appunti Anatomia VeterinariaDocumento25 pagineAppunti Anatomia VeterinariasimoneNessuna valutazione finora
- S Bob Anatomia Pa To Logic ADocumento59 pagineS Bob Anatomia Pa To Logic ADaniel RoxNessuna valutazione finora
- Vetrini Di IstologiaDocumento51 pagineVetrini Di Istologiawankerowl100% (8)
- La Cura Dda CompressamenteDocumento185 pagineLa Cura Dda CompressamentefifiNessuna valutazione finora
- File Completo IstologiaDocumento56 pagineFile Completo IstologiaSara Chiavacci100% (1)