Potrebbero piacerti anche
- 07 Testo Piante MedicinaliDocumento14 pagine07 Testo Piante MedicinaliMarro AlfredoNessuna valutazione finora
- Manuale Del MassaggioDocumento23 pagineManuale Del MassaggioGiorgio470% (2)
- Etnobotanica e Alimentazione Riflessioni Antropologiche Sull Utilizzo Delle Piante SpontaneeDocumento101 pagineEtnobotanica e Alimentazione Riflessioni Antropologiche Sull Utilizzo Delle Piante SpontaneefabriziozaraNessuna valutazione finora
- I FunghiDocumento274 pagineI FunghiAndrea GhidiniNessuna valutazione finora
- Scheda Fitoallimurgico E314 Parte1Documento7 pagineScheda Fitoallimurgico E314 Parte1caravindaNessuna valutazione finora
- FIBROMIALGIADocumento92 pagineFIBROMIALGIAlauretta100Nessuna valutazione finora
- FIBROMIALGIADocumento92 pagineFIBROMIALGIAlauretta100Nessuna valutazione finora
- Registri Akashici in Pratica!Documento15 pagineRegistri Akashici in Pratica!Malone Atropa100% (1)
- Specie Arbustive Macchia MediterraneaDocumento181 pagineSpecie Arbustive Macchia Mediterraneas_conoscenti8741100% (2)
- Nuove Letture Botaniche - Giovanni AliottaDocumento348 pagineNuove Letture Botaniche - Giovanni AliottaAnonymous pSCWlL3BdNessuna valutazione finora
- 1000 Piante Per Guarire Dal Cancro Senza Chemio Di G. NacciDocumento421 pagine1000 Piante Per Guarire Dal Cancro Senza Chemio Di G. Nacciprincy2006Nessuna valutazione finora
- Francesca Pegozzo - Senso Di ColpaDocumento22 pagineFrancesca Pegozzo - Senso Di ColpaMmeSilviaNessuna valutazione finora
- Guida Flora Alpi CarnicheDocumento508 pagineGuida Flora Alpi CarnicheGaia DanielaNessuna valutazione finora
- Massaggio SportivoDocumento2 pagineMassaggio Sportivolauretta100Nessuna valutazione finora
- Guida Per Riconoscere 50 Alberi Del VenetoDocumento129 pagineGuida Per Riconoscere 50 Alberi Del VenetocactusmtbNessuna valutazione finora
- Terapia ForestaleDocumento128 pagineTerapia ForestaleGovinda ChiesaNessuna valutazione finora
- Pignatti Et Al 2003 PDFDocumento328 paginePignatti Et Al 2003 PDFdemilich100% (1)
- Dizionario BotanicoDocumento241 pagineDizionario BotanicoDouglas Malone100% (2)
- Guida Pratica Alla Flora Della Sardegna PDFDocumento622 pagineGuida Pratica Alla Flora Della Sardegna PDFAlessandro Spanu100% (1)
- Roy MartinaDocumento35 pagineRoy MartinaPaola BosiNessuna valutazione finora
- Roy MartinaDocumento35 pagineRoy MartinaPaola BosiNessuna valutazione finora
- Roy MartinaDocumento35 pagineRoy MartinaPaola BosiNessuna valutazione finora
- Natura 102-1 2012Documento187 pagineNatura 102-1 2012Panda666Nessuna valutazione finora
- Il Nespolo Del Giappone, Origine e Aspetti Botanici PDFDocumento65 pagineIl Nespolo Del Giappone, Origine e Aspetti Botanici PDFMarcoNessuna valutazione finora
- Atlante Dei Funghi Commestibili Della BasilicataDocumento348 pagineAtlante Dei Funghi Commestibili Della BasilicataAngelo FigundioNessuna valutazione finora
- 1 PBDocumento10 pagine1 PBrhrwthrtrNessuna valutazione finora
- Centro Micologico Bollettino 2021 WebDocumento52 pagineCentro Micologico Bollettino 2021 WebMichal MikšíkNessuna valutazione finora
- Atlante Della Biodiversit Del Parco Adda SudDocumento290 pagineAtlante Della Biodiversit Del Parco Adda SudsalzilloNessuna valutazione finora
- La FarfallaDocumento43 pagineLa FarfallaMarta PalenzonaNessuna valutazione finora
- Sarnari - 2007 - Monografia Illustrata Del Genere Russula in EuropaDocumento769 pagineSarnari - 2007 - Monografia Illustrata Del Genere Russula in EuropaElías Peña VeraNessuna valutazione finora
- Sarnari - 2007 - Monografia Illustrata Del Genere Russula in EuropaDocumento802 pagineSarnari - 2007 - Monografia Illustrata Del Genere Russula in EuropaElías Peña VeraNessuna valutazione finora
- Variazione Degli Animali e Delle Piante Allo Stato Domestico - Charles DarwinDocumento522 pagineVariazione Degli Animali e Delle Piante Allo Stato Domestico - Charles DarwingbhendrixNessuna valutazione finora
- 089-197 Atti Botanica Farmaceutica Ed EtnobotanicaDocumento109 pagine089-197 Atti Botanica Farmaceutica Ed EtnobotanicacoloromaneNessuna valutazione finora
- Parlatorea Vol 5Documento125 pagineParlatorea Vol 5nicungNessuna valutazione finora
- Inula ViscosaDocumento122 pagineInula ViscosaRomeo ŠainNessuna valutazione finora
- Fauna Della Val RosandraDocumento117 pagineFauna Della Val RosandraDiego BerdonNessuna valutazione finora
- 1 IntroduzioneDocumento40 pagine1 IntroduzioneAngelo TranoNessuna valutazione finora
- ActaSucculenta 1 1 2013 ITDocumento94 pagineActaSucculenta 1 1 2013 ITlauretta100Nessuna valutazione finora
- Gli Enigmi Della Scienza Agosto 2017Documento69 pagineGli Enigmi Della Scienza Agosto 2017carlos_pro_5Nessuna valutazione finora
- Gli Effetti Della Fecondazione Incrociata - Charle DarwinDocumento228 pagineGli Effetti Della Fecondazione Incrociata - Charle DarwingbhendrixNessuna valutazione finora
- AtlanteAnfibi GrossetoDocumento116 pagineAtlanteAnfibi GrossetoAlex VrankenNessuna valutazione finora
- Actaplantarum Notes Aprile 2013Documento144 pagineActaplantarum Notes Aprile 2013mauri751Nessuna valutazione finora
- FAN LibroDocumento630 pagineFAN LibroJosue Laime QuiqueNessuna valutazione finora
- Dizionario Delle Scienze Naturali 1834 PG 615 Volume 6stoDocumento615 pagineDizionario Delle Scienze Naturali 1834 PG 615 Volume 6stoCraccoDrappoTafferoNessuna valutazione finora
- Flute TecniqueDocumento2 pagineFlute TecniqueIreneNessuna valutazione finora
- Piante Grasse - GiuntiDocumento130 paginePiante Grasse - Giuntimartina palumboNessuna valutazione finora
- EvoluzionismoDocumento21 pagineEvoluzionismoSilvia FermiNessuna valutazione finora
- I Macrolepidotteri ItalianiDocumento1.051 pagineI Macrolepidotteri ItalianiFrancesco100% (4)
- Botanica Pianta & Inquinamento AriaDocumento248 pagineBotanica Pianta & Inquinamento AriaRanieri FrattaroloNessuna valutazione finora
- Botanica SistematicaDocumento13 pagineBotanica SistematicakevinNessuna valutazione finora
- Il Fiore Perduto. La Cultura Del Papaver Somniferum Nell'area Del MediterraneoDocumento109 pagineIl Fiore Perduto. La Cultura Del Papaver Somniferum Nell'area Del MediterraneoMaurizio BelperioNessuna valutazione finora
- Gli Anfipodi Caprellidi Presenti Sulle Strutture Galleggianti Di Un Impianto Di Mitilicoltura OffshoreDocumento22 pagineGli Anfipodi Caprellidi Presenti Sulle Strutture Galleggianti Di Un Impianto Di Mitilicoltura OffshoreAlessandro NeriNessuna valutazione finora
- EchinocereusDocumento113 pagineEchinocereusEDESMO13100% (1)
- Il Senso Della ViteDocumento5 pagineIl Senso Della ViteAndrea BiffiNessuna valutazione finora
- Miglietta & Belmonte, 1995 - Bibliografia P. ParenzanDocumento21 pagineMiglietta & Belmonte, 1995 - Bibliografia P. ParenzanGabriele MacrìNessuna valutazione finora
- L'insediamento Nei Nidi Artificiali Ed Il Ciclo Biologico Degli Uccelli Utili in FrutticolturaDocumento4 pagineL'insediamento Nei Nidi Artificiali Ed Il Ciclo Biologico Degli Uccelli Utili in FrutticolturaPietro Lorenzo PavanNessuna valutazione finora
- Breve Guida Alla Preparazione Dell'erbario (Piante Vascolari)Documento6 pagineBreve Guida Alla Preparazione Dell'erbario (Piante Vascolari)koxxoNessuna valutazione finora
- Alberi VenetiDocumento52 pagineAlberi VenetiocoxodoNessuna valutazione finora
- Aquariophylia Nr.9 Ottobre 2011Documento120 pagineAquariophylia Nr.9 Ottobre 2011vincenzo_vincenzo_96Nessuna valutazione finora
- Instapaper - Manuale Italiano Degli Habitat Della Rete Natura 2000Documento9 pagineInstapaper - Manuale Italiano Degli Habitat Della Rete Natura 2000Francesco BonelliNessuna valutazione finora
- La Moretta GrigiaDocumento18 pagineLa Moretta GrigiasalzilloNessuna valutazione finora
- Cs. Naturales (Marzo-Abril) 2024Documento12 pagineCs. Naturales (Marzo-Abril) 2024RutkeviajesNessuna valutazione finora
- Due Nuovi Ibridi Di Orchidee Del Salento.Documento2 pagineDue Nuovi Ibridi Di Orchidee Del Salento.Legambiente SalveNessuna valutazione finora
- Paleontologia - Geologia - Botanica - Zoologia - Storia e Filosofia Della Scienza Volume 9. Con Il Patrocinio Di - Comune Di PiacenzaDocumento206 paginePaleontologia - Geologia - Botanica - Zoologia - Storia e Filosofia Della Scienza Volume 9. Con Il Patrocinio Di - Comune Di PiacenzaGianluca BoninsegniNessuna valutazione finora
- La Flora Delle Groane Novità e ConfermeDocumento31 pagineLa Flora Delle Groane Novità e Confermelucag2008Nessuna valutazione finora
- Il Massaggio e Le Sue TecnicheDocumento24 pagineIl Massaggio e Le Sue Tecnichelauretta100Nessuna valutazione finora
- Piante GrasseDocumento5 paginePiante Grasselauretta100Nessuna valutazione finora
- Piante Da InternoDocumento10 paginePiante Da InternovaleNessuna valutazione finora
- FLORENTERDocumento1 paginaFLORENTERlauretta100Nessuna valutazione finora
- Le Piante Grasse o SucculenteDocumento45 pagineLe Piante Grasse o Succulentelauretta100Nessuna valutazione finora
- OsteonobrandDocumento50 pagineOsteonobrandlauretta100Nessuna valutazione finora
- Il Grande Libro Dei Cactus e Delle Piante Grasse EstrattoDocumento19 pagineIl Grande Libro Dei Cactus e Delle Piante Grasse EstrattoLucia DiaferioNessuna valutazione finora
- MassaggioDocumento4 pagineMassaggiolauretta100Nessuna valutazione finora
- Acqua Oligominerale "Ielo" in Diuresi: Studio Caso-ControlloDocumento7 pagineAcqua Oligominerale "Ielo" in Diuresi: Studio Caso-Controllolauretta100Nessuna valutazione finora
- Massaggio SportivoDocumento8 pagineMassaggio Sportivolauretta100Nessuna valutazione finora
- Massaggio SportivoDocumento8 pagineMassaggio Sportivolauretta100Nessuna valutazione finora
- 2 - Alimentazione e OsteoporosiDocumento3 pagine2 - Alimentazione e Osteoporosilauretta100Nessuna valutazione finora
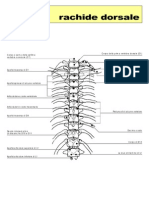
- Rachide DorsaleDocumento3 pagineRachide Dorsalelauretta100Nessuna valutazione finora
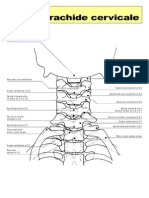
- Rachide CervicaleDocumento7 pagineRachide Cervicalelauretta100Nessuna valutazione finora
- Rachide DorsaleDocumento3 pagineRachide Dorsalelauretta100Nessuna valutazione finora
- Reflusso GastroesofageoDocumento6 pagineReflusso Gastroesofageolauretta100Nessuna valutazione finora
- Rachide CervicaleDocumento7 pagineRachide Cervicalelauretta100Nessuna valutazione finora
- UntitledDocumento2 pagineUntitledlauretta100Nessuna valutazione finora
- Documento FinaleDocumento68 pagineDocumento FinaleAnnalisa CangemiNessuna valutazione finora
- Normativa Illuminazione StradaleDocumento8 pagineNormativa Illuminazione StradaleLaura BiasiNessuna valutazione finora
- Controventi Instabilita Impedita ItaDocumento10 pagineControventi Instabilita Impedita Itaangelo_caroneNessuna valutazione finora
- 16 - Il Metodo Della Doppia Integrazione, o Di Saviotti PDFDocumento17 pagine16 - Il Metodo Della Doppia Integrazione, o Di Saviotti PDFPierpaolo ViggianoNessuna valutazione finora