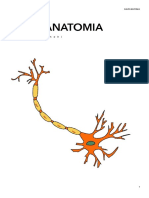
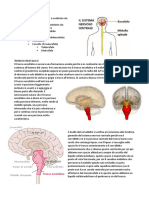
Potrebbero piacerti anche
- Il Cervello EmotivoDocumento43 pagineIl Cervello Emotivoluciano_tronchinNessuna valutazione finora
- Sul Sentiero I – Dalla “divina inquietudine” alla GioiaDa EverandSul Sentiero I – Dalla “divina inquietudine” alla GioiaNessuna valutazione finora
- Anima 18Documento16 pagineAnima 18zapremNessuna valutazione finora
- La casa che vorrei: Feng Shui, campane tibetane, oli essenziali e armonia interioreDa EverandLa casa che vorrei: Feng Shui, campane tibetane, oli essenziali e armonia interioreNessuna valutazione finora
- Il supernormale - Introduzione critica alla scienza psichicaDa EverandIl supernormale - Introduzione critica alla scienza psichicaNessuna valutazione finora
- 23 DiscepoloDocumento44 pagine23 Discepoloeternity21Nessuna valutazione finora
- "L'albero Del Giardino Posto Ad Oriente - La CabalaDocumento4 pagine"L'albero Del Giardino Posto Ad Oriente - La CabalaparisipromoNessuna valutazione finora
- Cortocircuito e tradimento nell'abuso sessuale infantileDa EverandCortocircuito e tradimento nell'abuso sessuale infantileNessuna valutazione finora
- Kirohermetica: La mano e il mito nella via alchemica occidentaleDa EverandKirohermetica: La mano e il mito nella via alchemica occidentaleNessuna valutazione finora
- Meditazione Guidata Per La Paura, Il Ripensamento E Le Preoccupazioni: Lascia Andare Via La Paura, I Troppi Pensieri E Lo Stress Mentre Dormi Meglio E Più A FondoDa EverandMeditazione Guidata Per La Paura, Il Ripensamento E Le Preoccupazioni: Lascia Andare Via La Paura, I Troppi Pensieri E Lo Stress Mentre Dormi Meglio E Più A FondoNessuna valutazione finora
- La rivoluzionaria teoria delle Intelligenze Multiple: Alcune valutazioni criticheDa EverandLa rivoluzionaria teoria delle Intelligenze Multiple: Alcune valutazioni criticheNessuna valutazione finora
- Cos È L A KabbbalahDocumento10 pagineCos È L A KabbbalahMassimo ANessuna valutazione finora
- Le nuove scienze e le antiche filosofie: Fisica quantistica, epigenetica, noetica in accordo con le antiche filosofie e religiosità sapienzaliDa EverandLe nuove scienze e le antiche filosofie: Fisica quantistica, epigenetica, noetica in accordo con le antiche filosofie e religiosità sapienzaliNessuna valutazione finora
- I Misteri EleusiniDocumento8 pagineI Misteri EleusiniMondo Comune100% (1)
- La Ghiandola Che Attiva La KundaliniDocumento2 pagineLa Ghiandola Che Attiva La Kundaliniamasino1Nessuna valutazione finora
- Abraxas N° 0Documento42 pagineAbraxas N° 0juanmatus2011Nessuna valutazione finora
- IL POTERE DELLA SEMPLICITA' Manuale di esercizi per l'autorealizzazioneDa EverandIL POTERE DELLA SEMPLICITA' Manuale di esercizi per l'autorealizzazioneNessuna valutazione finora
- Il potere della mente subconscia: Come mettere a frutto i nostri potenziali nascosti per raggiungere gli obiettivi e vivere una vita felice ed appaganteDa EverandIl potere della mente subconscia: Come mettere a frutto i nostri potenziali nascosti per raggiungere gli obiettivi e vivere una vita felice ed appaganteNessuna valutazione finora
- Jung e LastrologiaDocumento8 pagineJung e Lastrologiadavide_negri_4Nessuna valutazione finora
- Capire il Karma: Cinque conferenze tenute a Berlino e Stoccarda nel 1912Da EverandCapire il Karma: Cinque conferenze tenute a Berlino e Stoccarda nel 1912Nessuna valutazione finora
- E se non fosse depressione?: Un approccio innovativo e la soluzione definitivaDa EverandE se non fosse depressione?: Un approccio innovativo e la soluzione definitivaNessuna valutazione finora
- Cancro - una malattia curabile: Nuove scoperte in medicinaDa EverandCancro - una malattia curabile: Nuove scoperte in medicinaNessuna valutazione finora
- La Sindrome FrontaleDocumento7 pagineLa Sindrome FrontaleNeuroscienze.netNessuna valutazione finora
- Mater TenebrarumDocumento86 pagineMater TenebrarumVittorio BaccelliNessuna valutazione finora
- Segretezza Simboli ReikiDocumento14 pagineSegretezza Simboli Reikipaolo continiNessuna valutazione finora
- Dall'amore cieco all'amore consapevole. Relazione tra la pedagogia e le costellazioni di Bert HellingerDa EverandDall'amore cieco all'amore consapevole. Relazione tra la pedagogia e le costellazioni di Bert HellingerNessuna valutazione finora
- Ecce Quam Bonum 25Documento41 pagineEcce Quam Bonum 25Filippo GotiNessuna valutazione finora
- Entanglement e sincronicità. Campi di forza, non-località, percezioni extrasensoriali. Le sorprendenti proprietà della fisica quantistica.Da EverandEntanglement e sincronicità. Campi di forza, non-località, percezioni extrasensoriali. Le sorprendenti proprietà della fisica quantistica.Nessuna valutazione finora
- Nati per essere liberi: Consigli e pratiche per portare armonia in sé e nelle relazioni affettive attraverso il metodo delle costellazioni dell’amoreDa EverandNati per essere liberi: Consigli e pratiche per portare armonia in sé e nelle relazioni affettive attraverso il metodo delle costellazioni dell’amoreNessuna valutazione finora
- I Segreti dell'Alchimia: Come trasformare i sogni in realtàDa EverandI Segreti dell'Alchimia: Come trasformare i sogni in realtàNessuna valutazione finora
- Frequenze Cura ProblemiDocumento2 pagineFrequenze Cura ProblemiSkanda Lu Muruguan NooneNessuna valutazione finora
- Social cognition e aggressività. Disamina del modello socio-cognitivo di Kenneth A. DodgeDa EverandSocial cognition e aggressività. Disamina del modello socio-cognitivo di Kenneth A. DodgeNessuna valutazione finora
- Astrologia Evolutiva: Il Viaggio dell'Anima attraverso gli Stati della CoscienzaDa EverandAstrologia Evolutiva: Il Viaggio dell'Anima attraverso gli Stati della CoscienzaNessuna valutazione finora
- Viva il Movimento. 400 Giochi per bambini dall'età prescolare alle soglie dell'adolescenzaDa EverandViva il Movimento. 400 Giochi per bambini dall'età prescolare alle soglie dell'adolescenzaNessuna valutazione finora
- Reiki Shamanico: Modi Ampliati Di Lavorare Con L'energia Della Forza Vitale UniversaleDa EverandReiki Shamanico: Modi Ampliati Di Lavorare Con L'energia Della Forza Vitale UniversaleNessuna valutazione finora
- Lex Aurea 56Documento57 pagineLex Aurea 56Filippo GotiNessuna valutazione finora
- Aspetti Della Personalita JUNGDocumento336 pagineAspetti Della Personalita JUNGpatriciacostellazionifamiliariNessuna valutazione finora
- Alla ricerca delle ultime cose: vie spirituali e spunti escatologiciDa EverandAlla ricerca delle ultime cose: vie spirituali e spunti escatologiciNessuna valutazione finora
- Il Mito di Dracula: dall’oscurità delle origini, ai meandri dell’inconscio, al buio delle sale cinematograficheDa EverandIl Mito di Dracula: dall’oscurità delle origini, ai meandri dell’inconscio, al buio delle sale cinematograficheNessuna valutazione finora
- Il Piano astrale La vita che ci attende: alla luce dell’Insegnamento del Cerchio Firenze 77Da EverandIl Piano astrale La vita che ci attende: alla luce dell’Insegnamento del Cerchio Firenze 77Nessuna valutazione finora
- Anime fiamme gemelle: Riti per incontrare l’altra metà della melaDa EverandAnime fiamme gemelle: Riti per incontrare l’altra metà della melaNessuna valutazione finora
- Meditazione Da Fare Il 12Documento8 pagineMeditazione Da Fare Il 12Vesna MladjenovicNessuna valutazione finora
- NEUROANATOMIADocumento172 pagineNEUROANATOMIAFrancesco LanzaNessuna valutazione finora
- NeurologiaDocumento50 pagineNeurologiaSalvatore FerraroNessuna valutazione finora
- Ortognatodonzia II File UnicoDocumento94 pagineOrtognatodonzia II File UnicoRoberta BaratteroNessuna valutazione finora
- Anatomia 2 (Denise) AppuntiunipiDocumento158 pagineAnatomia 2 (Denise) AppuntiunipinuunumclNessuna valutazione finora
- CircolatorioDocumento41 pagineCircolatorioRiccardo Mason TroiaNessuna valutazione finora
- File Unico SMC4 Definitivo-243-311Documento69 pagineFile Unico SMC4 Definitivo-243-311CeceNessuna valutazione finora
- Sistema NervosoDocumento40 pagineSistema NervosoturchettinoNessuna valutazione finora
- Costellazioni Del CervellettoDocumento8 pagineCostellazioni Del CervellettoAnonymous p6x3UDNessuna valutazione finora
- Riassunto LibroDocumento33 pagineRiassunto LibromarinaNessuna valutazione finora
- Il Sistema Nervoso Centrale È Costituito DaDocumento5 pagineIl Sistema Nervoso Centrale È Costituito DaFrancesca CancelliereNessuna valutazione finora
- Tirocinio (E.O. Neurologico) 5Documento22 pagineTirocinio (E.O. Neurologico) 5Matteo FerrariNessuna valutazione finora
- TTD - TMU - Lezioni I - II - IIIDocumento17 pagineTTD - TMU - Lezioni I - II - IIIMaegazine EnizageamNessuna valutazione finora
- Espressione CorporeaDocumento184 pagineEspressione CorporeaBenedetto RutiloNessuna valutazione finora
- 21-04-08 Vie Della Sensibilit+á GeneraleDocumento7 pagine21-04-08 Vie Della Sensibilit+á GeneraleGiovanni VescovoNessuna valutazione finora
- Nuova Medicina ITA - Le 5 Leggi Di Hamer - FracitaDocumento4 pagineNuova Medicina ITA - Le 5 Leggi Di Hamer - FracitaPaolo VirgataNessuna valutazione finora
- Vie NervoseDocumento5 pagineVie Nervosexxx9090Nessuna valutazione finora
- Neurologia - RiassuntoDocumento38 pagineNeurologia - Riassuntoapi-3863584100% (3)
- OculomotricitàDocumento55 pagineOculomotricitàCipriano D'AmbrosioNessuna valutazione finora
- La Mia Storia Rev1-09Documento152 pagineLa Mia Storia Rev1-09Luisella BenedettiniNessuna valutazione finora
- Tabella Vie SNCDocumento3 pagineTabella Vie SNCLeonardo Pzl100% (1)
- NeurologiaDocumento226 pagineNeurologiaΑντώνιος Χατζηγεωργίου50% (2)
- Anatomia Del Sistema Nervoso Centrale - Schemi VisiviDocumento58 pagineAnatomia Del Sistema Nervoso Centrale - Schemi VisiviClara ScopellitiNessuna valutazione finora
- Facilitati Corpo UmDocumento26 pagineFacilitati Corpo Umsuperbi088 • kanekiNessuna valutazione finora
- Fisiologia Dell' Esercizio e Dello SportDocumento65 pagineFisiologia Dell' Esercizio e Dello SportDaniele Nervi33% (3)
- Neuroscience: Science of The Brain in ItalianDocumento60 pagineNeuroscience: Science of The Brain in ItalianInternational Brain Research Organization100% (1)
- Corpo UmanoDocumento60 pagineCorpo Umanozastekani mozak100% (2)
- Sistema NervosoDocumento22 pagineSistema NervosoLuca PanealboNessuna valutazione finora
- Sistema LimbicoDocumento13 pagineSistema LimbicoCarolina ArecaceaeNessuna valutazione finora
- Il CervellettoDocumento15 pagineIl CervellettoGiancarlo AttivissimoNessuna valutazione finora