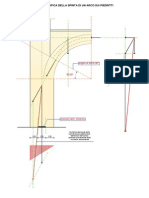
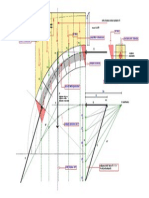
Potrebbero piacerti anche
- Stabil Arco 2Documento1 paginaStabil Arco 2savaziNessuna valutazione finora
- Mat5b7 10-11Documento2 pagineMat5b7 10-11savaziNessuna valutazione finora
- Mat5b3 10-11Documento1 paginaMat5b3 10-11savaziNessuna valutazione finora
- Mat5b2 09-10Documento1 paginaMat5b2 09-10savaziNessuna valutazione finora
- Mat5b8 09-10Documento2 pagineMat5b8 09-10savazi0% (1)
- Matematica Geom SolidaDocumento7 pagineMatematica Geom SolidasavaziNessuna valutazione finora
- Stabil Dell'arco1Documento1 paginaStabil Dell'arco1savaziNessuna valutazione finora
- Mat5b2 10-11Documento1 paginaMat5b2 10-11savaziNessuna valutazione finora
- Mat5b7 11-12Documento2 pagineMat5b7 11-12savaziNessuna valutazione finora
- Mat5b5 11-12Documento1 paginaMat5b5 11-12savaziNessuna valutazione finora
- Mat5b1 10-11Documento1 paginaMat5b1 10-11savaziNessuna valutazione finora
- Mat5b6 11-12Documento1 paginaMat5b6 11-12savaziNessuna valutazione finora
- Mat5b8 11-12Documento2 pagineMat5b8 11-12savaziNessuna valutazione finora
- Mat5b2 12-13Documento1 paginaMat5b2 12-13savaziNessuna valutazione finora
- Mat5a3 12-13Documento1 paginaMat5a3 12-13savaziNessuna valutazione finora
- Mat5b5 12-13Documento1 paginaMat5b5 12-13savaziNessuna valutazione finora
- Mat5ab7 12-13Documento2 pagineMat5ab7 12-13savaziNessuna valutazione finora
- Mat5a6 12-13Documento1 paginaMat5a6 12-13savaziNessuna valutazione finora
- Mat5b2 12-13Documento1 paginaMat5b2 12-13savaziNessuna valutazione finora
- Mat5b1 12-13Documento1 paginaMat5b1 12-13savaziNessuna valutazione finora
- Mat5a5 12-13Documento1 paginaMat5a5 12-13savaziNessuna valutazione finora
- Mat5a2 12-13Documento1 paginaMat5a2 12-13savaziNessuna valutazione finora
- Mat5a4 12-13Documento1 paginaMat5a4 12-13savaziNessuna valutazione finora
- Mat5a1 12-13Documento1 paginaMat5a1 12-13savaziNessuna valutazione finora
- Mat5b6 11-12Documento1 paginaMat5b6 11-12savaziNessuna valutazione finora
- Ese 07 8Documento2 pagineEse 07 8savaziNessuna valutazione finora
- Mat5b6 11-12Documento1 paginaMat5b6 11-12savaziNessuna valutazione finora
- FunzioniDocumento5 pagineFunzionisavaziNessuna valutazione finora
- Ese 07 7Documento2 pagineEse 07 7savaziNessuna valutazione finora
- Ese 07 9Documento2 pagineEse 07 9savaziNessuna valutazione finora
- Domande A Risposta Multipla LipidiDocumento5 pagineDomande A Risposta Multipla Lipidibiagio castronovoNessuna valutazione finora
- 10 - MMS - Protocollo 1000+ (Più) in ITALIANODocumento3 pagine10 - MMS - Protocollo 1000+ (Più) in ITALIANOMauro Lore100% (1)
- 18 Alcheni-Enoli PDFDocumento20 pagine18 Alcheni-Enoli PDFrodrigue yanNessuna valutazione finora
- Tabella Proprietà Composti OrganiciDocumento8 pagineTabella Proprietà Composti OrganiciLovelyJadeNessuna valutazione finora
- Esercizi Chimica OrganicaDocumento108 pagineEsercizi Chimica OrganicaminchiazzurraNessuna valutazione finora
- ANTAGONISTI MUSCARINICI - ClassificazioneDocumento5 pagineANTAGONISTI MUSCARINICI - Classificazioneangelobasile88Nessuna valutazione finora