Potrebbero piacerti anche
- Channels, Carriers, and Pumps: An Introduction to Membrane TransportDa EverandChannels, Carriers, and Pumps: An Introduction to Membrane TransportNessuna valutazione finora
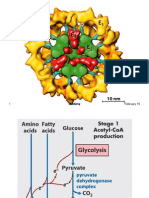
- PDH Complex and TCA CycleDocumento20 paginePDH Complex and TCA CycleDarrion LouisNessuna valutazione finora
- Biochemistry Saeed Oraby Part 3Documento276 pagineBiochemistry Saeed Oraby Part 3أسہہآمہهہ شہعہلآن100% (3)
- Respiratory Chain & Oxidative PhosphorylationDocumento57 pagineRespiratory Chain & Oxidative PhosphorylationHanifa AffianiNessuna valutazione finora
- Krebs Cycle: CHEM 160Documento47 pagineKrebs Cycle: CHEM 160Gianna Kristen MirandaNessuna valutazione finora
- Lehninger Ch26Documento81 pagineLehninger Ch26AMAN KUMAR SINGH100% (1)
- Multidimensional Analysis of Clinical Symptoms in Patients With Fabry's DiseaseDocumento12 pagineMultidimensional Analysis of Clinical Symptoms in Patients With Fabry's DiseaseOnon EssayedNessuna valutazione finora
- Biochemistry of Kidneys and UrineDocumento18 pagineBiochemistry of Kidneys and UrineAndrias PutriNessuna valutazione finora
- Biomolecules: Amino Acids, Peptides, and ProteinsDocumento55 pagineBiomolecules: Amino Acids, Peptides, and ProteinsBoooooooseNessuna valutazione finora
- CH 19 Integration of MetabolismDocumento63 pagineCH 19 Integration of Metabolismshee140802100% (1)
- Chapter 18 Lippincott BiochemistryDocumento62 pagineChapter 18 Lippincott BiochemistryMeysam SajjadiNessuna valutazione finora
- Metabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaDocumento60 pagineMetabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaAmanda PutriNessuna valutazione finora
- Integration of Metabolism: Dr. Farzana Hakim Assistant Professor Biochemistry DepartmentDocumento63 pagineIntegration of Metabolism: Dr. Farzana Hakim Assistant Professor Biochemistry DepartmentGriffinNessuna valutazione finora
- Mechanism of Action of NeurotransmitterDocumento10 pagineMechanism of Action of NeurotransmitterIrah FloresNessuna valutazione finora
- Adrenergic and Anti-Adrenergic DrugsDocumento54 pagineAdrenergic and Anti-Adrenergic DrugsChittaranjan Padhy100% (1)
- Carbohydrates MetabolismDocumento6 pagineCarbohydrates MetabolismAisha ShahfiqueeNessuna valutazione finora
- Biochemistry One LinersDocumento11 pagineBiochemistry One Linerssandeep MahapatraNessuna valutazione finora
- Glycogen MetabolismDocumento23 pagineGlycogen Metabolismessa100% (1)
- Biochem QbankDocumento16 pagineBiochem Qbank786waqar786Nessuna valutazione finora
- Biochem HomeworkDocumento13 pagineBiochem Homeworkfcukingfranztastik50% (2)
- Nucleotide Biosynthesis Lecture 2017sDocumento23 pagineNucleotide Biosynthesis Lecture 2017sSaad KazmiNessuna valutazione finora
- PDH Complex REVIEW: DR Navin SDocumento23 paginePDH Complex REVIEW: DR Navin Svaseem1981Nessuna valutazione finora
- S13.CHO .MetabolismDocumento18 pagineS13.CHO .MetabolismGhea Jovita SinagaNessuna valutazione finora
- MBBS MSK YR1 Muscles of Upper LimbDocumento52 pagineMBBS MSK YR1 Muscles of Upper Limbcharlesy TNessuna valutazione finora
- Metabolism Lippincott's Q&ADocumento86 pagineMetabolism Lippincott's Q&Alina amjadNessuna valutazione finora
- Biochemistry Notes For BoardsDocumento35 pagineBiochemistry Notes For BoardsUjjwal PyakurelNessuna valutazione finora
- PROTEIN METABOLISM Dea Farha Fira Darson FineDocumento44 paginePROTEIN METABOLISM Dea Farha Fira Darson FineFarhati MardhiyahNessuna valutazione finora
- 1 Red Blood Cells Anemia and PolycythemiaDocumento43 pagine1 Red Blood Cells Anemia and PolycythemiaGeevee Naganag VentulaNessuna valutazione finora
- Biochemistry-Exam 2 Heinz SchwarzkopfDocumento162 pagineBiochemistry-Exam 2 Heinz SchwarzkopfthomasNessuna valutazione finora
- Integration of Metabolism 2019Documento68 pagineIntegration of Metabolism 2019Cahyani Tiara Safitri100% (1)
- Glycogenolysis: BiochemistryDocumento6 pagineGlycogenolysis: BiochemistryManila MedNessuna valutazione finora
- The Fate of Metabolism Andmabolic PathwaysDocumento25 pagineThe Fate of Metabolism Andmabolic PathwaysXuân Vi100% (1)
- Lecture 6: Part 1 Oxidative Phosphorylation and ETC: Produced by Samira Aili Presented by Alvaro GarciaDocumento23 pagineLecture 6: Part 1 Oxidative Phosphorylation and ETC: Produced by Samira Aili Presented by Alvaro Garciakristal elias100% (1)
- Anatomy and Physiology Summary Body Fluids and BloodDocumento22 pagineAnatomy and Physiology Summary Body Fluids and BloodKelly Anne BrandNessuna valutazione finora
- Porphyrias and Errors in Heme Metabolism: Rakesh Sharma, PH.DDocumento33 paginePorphyrias and Errors in Heme Metabolism: Rakesh Sharma, PH.DProfessor Rakesh Sharma Biochemistry Lectures0% (1)
- Uronic Acid PathwayDocumento4 pagineUronic Acid PathwayAlen Arthur100% (2)
- Pyruvate Dehydrogenase LectureDocumento34 paginePyruvate Dehydrogenase LectureFerasKarajehNessuna valutazione finora
- Fed Fast CycleDocumento7 pagineFed Fast CycleZahra AlaradiNessuna valutazione finora
- The Action Potential: Page 1. IntroductionDocumento12 pagineThe Action Potential: Page 1. IntroductionSimón JimenezNessuna valutazione finora
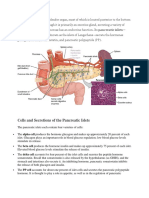
- Cells and Secretions of The Pancreatic IsletsDocumento4 pagineCells and Secretions of The Pancreatic IsletsSophia OcayNessuna valutazione finora
- Kreb CycleDocumento6 pagineKreb Cycle211BT014 Jeev Sheen JosephNessuna valutazione finora
- 8 Carbohydrate Metabolism PDFDocumento36 pagine8 Carbohydrate Metabolism PDFDayne Ocampo-SolimanNessuna valutazione finora
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocumento2 pagineHW - Carbohydrate Metabolism II & Lipid MetabolismyanNessuna valutazione finora
- Chapter 2 P-2 Enzyme-Inhibition 1Documento39 pagineChapter 2 P-2 Enzyme-Inhibition 1Raihan I. SakibNessuna valutazione finora
- Biochemistry Midterm 1 QuestionsDocumento2 pagineBiochemistry Midterm 1 Questionselfin_treeNessuna valutazione finora
- Psychology KMTCDocumento250 paginePsychology KMTCMercy AdeolaNessuna valutazione finora
- Chapter 23 Lippincott BiochemistryDocumento44 pagineChapter 23 Lippincott BiochemistryMeysam SajjadiNessuna valutazione finora
- l08 Metabolism in Liver 2 - Fatty Acid Metabolism (Mod)Documento21 paginel08 Metabolism in Liver 2 - Fatty Acid Metabolism (Mod)Swapnil100% (4)
- Urea CycleDocumento6 pagineUrea CycleFaradillah AnjaniNessuna valutazione finora
- Some Biochem MnemonicsDocumento8 pagineSome Biochem MnemonicsMikey PalominoNessuna valutazione finora
- Final Exam (Practice) KEYDocumento11 pagineFinal Exam (Practice) KEYNevenKnezevicNessuna valutazione finora
- Biochemistry of Digestive SystemDocumento55 pagineBiochemistry of Digestive SystemSyam UnhasNessuna valutazione finora
- Musculoskeletal PhysiologyDocumento5 pagineMusculoskeletal PhysiologyKhairul IkhwanNessuna valutazione finora
- Metabolism of Purine and PyrimidineDocumento56 pagineMetabolism of Purine and PyrimidineAboubakar Moalim Mahad moh'dNessuna valutazione finora
- Starch and Sucrose MetabolismDocumento41 pagineStarch and Sucrose MetabolismSanaur Rahman0% (1)
- Integration of Metabolism Integration of MetabolismDocumento10 pagineIntegration of Metabolism Integration of MetabolismEdison LucianoNessuna valutazione finora
- Biochemistry Saeed Oraby Part 1Documento248 pagineBiochemistry Saeed Oraby Part 1Go Hell100% (2)
- HMP Pathway PDFDocumento44 pagineHMP Pathway PDFHimanshu SharmaNessuna valutazione finora
- Biochemistry (GREEN)Documento18 pagineBiochemistry (GREEN)BRYAN BALDOMERONessuna valutazione finora
- The Autonomic Nervous System and Its Central ControlDocumento15 pagineThe Autonomic Nervous System and Its Central Controlpuchio100% (1)
- Bioenergetics and Biological Oxidation PDFDocumento50 pagineBioenergetics and Biological Oxidation PDFGEROME ABENILLA100% (1)
- ENZYMES - Overview and NomenclatureDocumento22 pagineENZYMES - Overview and NomenclatureSeania BuenaventuraNessuna valutazione finora
- LIF101 Mid Semester ExaminationDocumento6 pagineLIF101 Mid Semester ExaminationPiyush KumarNessuna valutazione finora
- The Importance of Energy Changes and Electron Transfer in MetabolismDocumento19 pagineThe Importance of Energy Changes and Electron Transfer in Metabolismslow_8Nessuna valutazione finora
- International Journal of Scientific and Innovative Research 2015 3 (1) P-ISSN 2347-2189, E - ISSN 2347-4971Documento172 pagineInternational Journal of Scientific and Innovative Research 2015 3 (1) P-ISSN 2347-2189, E - ISSN 2347-4971Dr.B.R.PandeyNessuna valutazione finora
- Vitamin b2 - RiboflavinDocumento21 pagineVitamin b2 - Riboflavinapi-388948078Nessuna valutazione finora
- CHAPTER 17 The Metabolism An OverviewDocumento10 pagineCHAPTER 17 The Metabolism An Overview楊畯凱Nessuna valutazione finora
- Topic: Co-Enzyme ChemistryDocumento12 pagineTopic: Co-Enzyme ChemistryAshutosh PrasadNessuna valutazione finora
- Quiz Lehninger CH 16 FinalDocumento15 pagineQuiz Lehninger CH 16 Finalasjdg100% (2)
- Part 7 Lecture MitochondriaDocumento13 paginePart 7 Lecture MitochondriaEric Jones UyNessuna valutazione finora
- VitaminsDocumento77 pagineVitaminsLoredana100% (1)
- BIOCHEMISTRY Lecture NotesDocumento7 pagineBIOCHEMISTRY Lecture NotesFaye AquinoNessuna valutazione finora
- Oxidation Reduction ReactionsDocumento23 pagineOxidation Reduction ReactionsRussNessuna valutazione finora
- Nitrogen Metabolism: Nitrate AssimilationDocumento3 pagineNitrogen Metabolism: Nitrate AssimilationSalsabila LuqyanaNessuna valutazione finora
- A Brief Review of How To Construct An Enzyme-Based H O Sensor Involved in NanomaterialsDocumento25 pagineA Brief Review of How To Construct An Enzyme-Based H O Sensor Involved in Nanomaterialskumara1986Nessuna valutazione finora
- EJCHEM Volume 65 Issue 8 Pages 43-55Documento13 pagineEJCHEM Volume 65 Issue 8 Pages 43-55فردوس يسNessuna valutazione finora
- Gim Lec Chapter 4Documento88 pagineGim Lec Chapter 4Paolo NaguitNessuna valutazione finora
- High-Dose Vitamin Therapy Stimulates Variant Enzymes With Decreased Coenzyme Binding AffinityDocumento43 pagineHigh-Dose Vitamin Therapy Stimulates Variant Enzymes With Decreased Coenzyme Binding AffinityytreffalNessuna valutazione finora
- Enzymes LectureDocumento16 pagineEnzymes Lectureglenn johnstonNessuna valutazione finora
- Engineering of HalogenasesDocumento123 pagineEngineering of HalogenasesNick PapasNessuna valutazione finora
- How Cells Harvest EnergyDocumento26 pagineHow Cells Harvest EnergyMarita YaghiNessuna valutazione finora
- Mil4ch32 (How Animals Harvest)Documento10 pagineMil4ch32 (How Animals Harvest)Muhammad AsadkhanNessuna valutazione finora
- Carbohydrate Metabolism 1Documento70 pagineCarbohydrate Metabolism 1nia adelleNessuna valutazione finora
- Chapter 6 McKee Enzyme KineticsDocumento79 pagineChapter 6 McKee Enzyme KineticsSuwahono, M.PdNessuna valutazione finora
- Screenshot 2023-03-28 at 11.36.42 AMDocumento48 pagineScreenshot 2023-03-28 at 11.36.42 AMSourayaNessuna valutazione finora
- Chapter9 CellularRespirationFermentation - Student - BIO21 - SMCDocumento40 pagineChapter9 CellularRespirationFermentation - Student - BIO21 - SMCSofiaNessuna valutazione finora
- Wardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test BankDocumento38 pagineWardlaws Perspectives in Nutrition A Functional Approach 1st Edition Byrd Bredbenner Test Bankedricduyenuc1uw100% (25)
- Exam 4 KEYDocumento12 pagineExam 4 KEYcwodNessuna valutazione finora
- Lecture 2PBEDocumento44 pagineLecture 2PBEJawadNessuna valutazione finora