Potrebbero piacerti anche
- Adventitious Root FR Deadline TOMORROWDocumento9 pagineAdventitious Root FR Deadline TOMORROWCherryAnnEspantoNessuna valutazione finora
- Translocation of SolutesDocumento22 pagineTranslocation of Solutesmanojtbgri5793Nessuna valutazione finora
- VATech WabagDocumento4 pagineVATech WabagPooja YamiNessuna valutazione finora
- Vinoxide HTC PDFDocumento5 pagineVinoxide HTC PDFTAN LI QINessuna valutazione finora
- Vitros Microslide Assay SummaryDocumento10 pagineVitros Microslide Assay SummaryMedLab Şamaxı KlinikasıNessuna valutazione finora
- Auxin: Greek Auxein, Meaning "To Increase" or "To Grow."Documento31 pagineAuxin: Greek Auxein, Meaning "To Increase" or "To Grow."gouthami shivaswamyNessuna valutazione finora
- Lecture On Plant HormonesDocumento9 pagineLecture On Plant HormonesPreen SanmukNessuna valutazione finora
- MBU Plant Hormones 22Documento41 pagineMBU Plant Hormones 22ketakeeNessuna valutazione finora
- Plant Growth RegulatorsDocumento55 paginePlant Growth RegulatorsXimena Libano0% (1)
- Plant HormoneDocumento31 paginePlant HormoneChristine De GuzmanNessuna valutazione finora
- Tackling Exam Questions - Plant Growth SubstancesDocumento3 pagineTackling Exam Questions - Plant Growth SubstancesomarNessuna valutazione finora
- 18BTE310T - Introduction To Phytohormones - Unit - IDocumento118 pagine18BTE310T - Introduction To Phytohormones - Unit - IAkhila SarvasuddiNessuna valutazione finora
- Planthormones 141025004114 Conversion Gate02 PDFDocumento75 paginePlanthormones 141025004114 Conversion Gate02 PDFSaiTimmarao100% (1)
- Respiration in PlantDocumento25 pagineRespiration in PlantFatimah AzzahrahNessuna valutazione finora
- Affect of Phytohormons in Leaf SenescenceDocumento29 pagineAffect of Phytohormons in Leaf Senescencemasoomi_farhad2030Nessuna valutazione finora
- What Is The 16S RRNA GeneDocumento3 pagineWhat Is The 16S RRNA GeneBenjamin UcichNessuna valutazione finora
- Biodegradable Polymers From Corn's StarchDocumento32 pagineBiodegradable Polymers From Corn's StarchMuhammad ArshadNessuna valutazione finora
- MGE Galaxy 3500 enDocumento48 pagineMGE Galaxy 3500 en11111Nessuna valutazione finora
- Sulphur CycleDocumento5 pagineSulphur CycleAkshat Khandelwal0% (1)
- Aglaonema MicropropagationDocumento5 pagineAglaonema MicropropagationsonNessuna valutazione finora
- In Vitro Propagation in Pteridophytes A ReviewDocumento7 pagineIn Vitro Propagation in Pteridophytes A Reviewathira vijayanNessuna valutazione finora
- Lesson 4.2. GlycoconjugatesDocumento4 pagineLesson 4.2. GlycoconjugatesGemma CabañasNessuna valutazione finora
- DNA Extraction LabDocumento2 pagineDNA Extraction LabNur'Izzah Juarah100% (1)
- An Analysis of Variation of The Aroma Gene in Rice (Oryza Sativa L. Subsp. Indica Kato) LandracesDocumento7 pagineAn Analysis of Variation of The Aroma Gene in Rice (Oryza Sativa L. Subsp. Indica Kato) LandracesTiaChakrabortyNessuna valutazione finora
- Phytohormone - Types and FunctionsDocumento23 paginePhytohormone - Types and FunctionsminiNessuna valutazione finora
- Importance of Plant Tissue CultureDocumento16 pagineImportance of Plant Tissue Culturealphabetagama713085100% (10)
- Use of Microbes in AgricultureDocumento4 pagineUse of Microbes in AgricultureAran MuthNessuna valutazione finora
- Molecular Systematics - David Hillis, Craig Moritz, Barbara MableDocumento676 pagineMolecular Systematics - David Hillis, Craig Moritz, Barbara MableFellipNessuna valutazione finora
- Nitrogen Fixation PDFDocumento9 pagineNitrogen Fixation PDFCaspiaNessuna valutazione finora
- Morphology of Root (By Hemraj Aryal)Documento23 pagineMorphology of Root (By Hemraj Aryal)sabita basnetNessuna valutazione finora
- Rice Industry in MalaysiaDocumento27 pagineRice Industry in MalaysiaYatt T PhrofileNessuna valutazione finora
- Enzyme Technology - CHAPLIN PDFDocumento316 pagineEnzyme Technology - CHAPLIN PDFSharan Gayathrinathan100% (1)
- Network Structure in Root, Stem, and LeafDocumento5 pagineNetwork Structure in Root, Stem, and LeafMarwana SuaibNessuna valutazione finora
- Practical 3 Botany DrawingDocumento7 paginePractical 3 Botany DrawingSleeping BeautyNessuna valutazione finora
- Cyanobacteria Economic ImportanceDocumento15 pagineCyanobacteria Economic ImportanceKumail IslamianNessuna valutazione finora
- Basic Principles and Protocol in Plant Tissue CultureDocumento17 pagineBasic Principles and Protocol in Plant Tissue CultureHelenChungNessuna valutazione finora
- Biomolecules and Cells:: Mr. Derrick Banda MSC, BSCDocumento69 pagineBiomolecules and Cells:: Mr. Derrick Banda MSC, BSCAmon Sangulube100% (1)
- Agricultural Practices of MalaysiaDocumento41 pagineAgricultural Practices of MalaysiaWan Shahrizal100% (2)
- Environmental Quality Act in MalaysiaDocumento18 pagineEnvironmental Quality Act in MalaysiaKALESMI A/P AMARALATHAN STUDENTNessuna valutazione finora
- Moisture Stress of PlantDocumento30 pagineMoisture Stress of PlantHassen100% (1)
- PHD Thesis. Cathrine H. IngvordsenDocumento187 paginePHD Thesis. Cathrine H. IngvordsenIngenioerenNessuna valutazione finora
- Papaya in VitroDocumento5 paginePapaya in VitroBiomontec Biomontec BiomontecNessuna valutazione finora
- PestsDocumento15 paginePestsRaquel DomingoNessuna valutazione finora
- Biodiversity in MalaysiaDocumento36 pagineBiodiversity in MalaysiaJu' NanieNessuna valutazione finora
- Lecture 32 PCR & DNA ExtractionDocumento25 pagineLecture 32 PCR & DNA ExtractionJenna ScheiblerNessuna valutazione finora
- PERKEMBANGAN KULTUR DAUN Aglaonema Sp. Var Siam Pearl, Aglaonema Sp. Var. Lady Valentin Dan Aglaonema Sp. Var. Lipstik DENGAN PERLAKUAN ZAT PENGATUR TUMBUH IAA DAN BAPDocumento7 paginePERKEMBANGAN KULTUR DAUN Aglaonema Sp. Var Siam Pearl, Aglaonema Sp. Var. Lady Valentin Dan Aglaonema Sp. Var. Lipstik DENGAN PERLAKUAN ZAT PENGATUR TUMBUH IAA DAN BAPHery PurnobasukiNessuna valutazione finora
- Nitrogen Cycling in EcosystemsDocumento7 pagineNitrogen Cycling in EcosystemsMark Abion ValladolidNessuna valutazione finora
- Fitohormon IntroDocumento27 pagineFitohormon Introvitri_indrianiNessuna valutazione finora
- PhotoperiodismDocumento14 paginePhotoperiodismArun Sreenivasan IyengarNessuna valutazione finora
- In Vitro Regeneration and Somatic Embryogenesis and Their Antimicrobial Studies in Selected Ornamental PlantsDocumento20 pagineIn Vitro Regeneration and Somatic Embryogenesis and Their Antimicrobial Studies in Selected Ornamental Plantsbakru248326Nessuna valutazione finora
- Plant Growth RegulatorsDocumento28 paginePlant Growth RegulatorsFaishol MuhammadNessuna valutazione finora
- Recovery Processes of Organic Acids From Fermentation Broths inDocumento8 pagineRecovery Processes of Organic Acids From Fermentation Broths inRocio ChongNessuna valutazione finora
- Abiotic and Biotic Stresses in Soybean Production: Soybean Production Volume 1Da EverandAbiotic and Biotic Stresses in Soybean Production: Soybean Production Volume 1Valutazione: 4 su 5 stelle4/5 (1)
- Breeding The Papaya - Carica Papaya PDFDocumento11 pagineBreeding The Papaya - Carica Papaya PDFchikkannaswamyNessuna valutazione finora
- Xylem and PhloemDocumento35 pagineXylem and PhloemAvanish PratapNessuna valutazione finora
- 00 Key For The Identification of Different BambooDocumento14 pagine00 Key For The Identification of Different BambooShanaia BualNessuna valutazione finora
- Phylogeny AnalysisDocumento49 paginePhylogeny AnalysisQuyen Nguyen Thi AnhNessuna valutazione finora
- Mechanism of Absorption of WaterDocumento6 pagineMechanism of Absorption of WaterketakeeNessuna valutazione finora
- Plant Growth - HormonesDocumento34 paginePlant Growth - HormonesKambaska BeheraNessuna valutazione finora
- Fifu Landskap MalaysiaDocumento108 pagineFifu Landskap MalaysiaMariam MrmNessuna valutazione finora
- The Action of Hormones in plants and invertebratesDa EverandThe Action of Hormones in plants and invertebratesKenneth ThimannNessuna valutazione finora
- Political Economic Social: GeographicDocumento2 paginePolitical Economic Social: GeographicamyNessuna valutazione finora
- Cell and DevDocumento3 pagineCell and DevamyNessuna valutazione finora
- BIOC2162 Tutorial #5 2019-20Documento1 paginaBIOC2162 Tutorial #5 2019-20amyNessuna valutazione finora
- Wiki Images, 2020Documento21 pagineWiki Images, 2020amyNessuna valutazione finora
- Jada Cumberbatch 816010507 BIOC2162 Secretory and Circulatory Systems Tutorial #2Documento14 pagineJada Cumberbatch 816010507 BIOC2162 Secretory and Circulatory Systems Tutorial #2amyNessuna valutazione finora
- Group 1 BIOC3062 - 2019 Virus Induced DiabetesDocumento46 pagineGroup 1 BIOC3062 - 2019 Virus Induced DiabetesamyNessuna valutazione finora
- Group 1 BIOC3062 - 2019 Virus Induced DiabetesDocumento46 pagineGroup 1 BIOC3062 - 2019 Virus Induced DiabetesamyNessuna valutazione finora
- Name: Jada CumberbatchDocumento6 pagineName: Jada CumberbatchamyNessuna valutazione finora
- ApoptosisDocumento36 pagineApoptosisamyNessuna valutazione finora
- Breeding Methods in Self-Pollinated CropsDocumento56 pagineBreeding Methods in Self-Pollinated Cropsamy100% (2)
- Alliance Connectivity Link According Con Flicts Example A - Reflection On Group WorkDocumento2 pagineAlliance Connectivity Link According Con Flicts Example A - Reflection On Group WorkamyNessuna valutazione finora
- Standardization 4Documento2 pagineStandardization 4amyNessuna valutazione finora
- PhytochromeDocumento27 paginePhytochromeamyNessuna valutazione finora
- Learning Objectives Cell Membrane and Ion Uptake-2Documento2 pagineLearning Objectives Cell Membrane and Ion Uptake-2amyNessuna valutazione finora
- Genetics Tutorial 5Documento5 pagineGenetics Tutorial 5amyNessuna valutazione finora
- Heralding The Doctrine of Double Effect Is The Concept of Abortion in Trinidad and TobagoDocumento1 paginaHeralding The Doctrine of Double Effect Is The Concept of Abortion in Trinidad and TobagoamyNessuna valutazione finora
- Wiki Images, 2020Documento21 pagineWiki Images, 2020amyNessuna valutazione finora
- Practical 3 & 4 - Germination, Mineral Nutrition and Hormone ResponseDocumento18 paginePractical 3 & 4 - Germination, Mineral Nutrition and Hormone ResponseamyNessuna valutazione finora
- Anova Test For Plants Lab 3&4Documento2 pagineAnova Test For Plants Lab 3&4amyNessuna valutazione finora
- GapsDocumento2 pagineGapsamyNessuna valutazione finora
- Factors Affecting Bimodal Respiration in The Catfish Corydoras AeneusDocumento2 pagineFactors Affecting Bimodal Respiration in The Catfish Corydoras AeneusamyNessuna valutazione finora
- NAME1Documento13 pagineNAME1amyNessuna valutazione finora
- Biol 2867 Lab 1 GraphsDocumento4 pagineBiol 2867 Lab 1 GraphsamyNessuna valutazione finora
- BIOL3769 Tutorial Question-2Documento1 paginaBIOL3769 Tutorial Question-2amyNessuna valutazione finora
- Tows Matrix - 3Documento2 pagineTows Matrix - 3amyNessuna valutazione finora
- Tutorial Worksheet #7 ETC #2 2019-20Documento1 paginaTutorial Worksheet #7 ETC #2 2019-20amyNessuna valutazione finora
- AutoRecovery Save of Business Strategy Assignment - 4Documento10 pagineAutoRecovery Save of Business Strategy Assignment - 4amyNessuna valutazione finora
- Photosynthesis Is An Endothermic Reaction That Occurs On The Thylakoid of The Chloroplasts in PlantsDocumento4 paginePhotosynthesis Is An Endothermic Reaction That Occurs On The Thylakoid of The Chloroplasts in PlantsamyNessuna valutazione finora
- Name: Jada CumberbatchDocumento6 pagineName: Jada CumberbatchamyNessuna valutazione finora
- Chemical Mediators of InflammationDocumento46 pagineChemical Mediators of Inflammationroohi khanNessuna valutazione finora
- Human Biology: Unit: 4HB0 Paper: 01Documento36 pagineHuman Biology: Unit: 4HB0 Paper: 01Joseph LAU [11D]Nessuna valutazione finora
- 1rki Physical AssesmentDocumento18 pagine1rki Physical AssesmentIrfan NasrullahNessuna valutazione finora
- Animal Tissue NotesDocumento20 pagineAnimal Tissue NotesSahejNessuna valutazione finora
- Negative Feedback Mechanisms: Lesson 15.2Documento20 pagineNegative Feedback Mechanisms: Lesson 15.2Jazmine MariezNessuna valutazione finora
- Physical Education XiDocumento59 paginePhysical Education XiAkshitNessuna valutazione finora
- Hypertension CASE REPORTDocumento57 pagineHypertension CASE REPORTJulienne Sanchez-SalazarNessuna valutazione finora
- NCMB314 Lec PrelimDocumento25 pagineNCMB314 Lec PrelimRen Mark CanlasNessuna valutazione finora
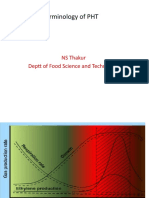
- Terminology of PHT-1Documento11 pagineTerminology of PHT-1Hebilstone SyniaNessuna valutazione finora
- Lipid MetabolismDocumento25 pagineLipid Metabolismബാദ്ഷാ ഖാൻNessuna valutazione finora
- X Study Material Final ModifiedDocumento317 pagineX Study Material Final Modifieddevisree100% (1)
- Answers To Eocqs: Notes About Mark SchemesDocumento3 pagineAnswers To Eocqs: Notes About Mark SchemesAayush GauchanNessuna valutazione finora
- Diagnosis and Treatment of Primary AldosteronismDocumento17 pagineDiagnosis and Treatment of Primary AldosteronismcastillojessNessuna valutazione finora
- Uti Case StudyDocumento11 pagineUti Case StudyJa MereNessuna valutazione finora
- Musculature and Fascia: Station 1: The Anterior Abdominal WallDocumento10 pagineMusculature and Fascia: Station 1: The Anterior Abdominal WallAlia SahniNessuna valutazione finora
- 1-Introduction To PhysiologyDocumento41 pagine1-Introduction To Physiologyfatimahdua572Nessuna valutazione finora
- Gnaps: Lomerulo Efritis KUT Asca TreptokokusDocumento32 pagineGnaps: Lomerulo Efritis KUT Asca TreptokokusRamsyifa VirzanisdaNessuna valutazione finora
- Concise Atlas of Basic HemopoiesisDocumento14 pagineConcise Atlas of Basic HemopoiesisdynndgrNessuna valutazione finora
- Life Sciences P1 Eng MemoDocumento12 pagineLife Sciences P1 Eng MemoabingdadeNessuna valutazione finora
- Introduction To Medical Mycology 2023Documento54 pagineIntroduction To Medical Mycology 2023Samuel fikaduNessuna valutazione finora
- Biology 11Th Edition Raven Test Bank Full Chapter PDFDocumento36 pagineBiology 11Th Edition Raven Test Bank Full Chapter PDFlewis.barnes1000100% (12)
- Development Psychology - Chapter 4 (Santrock)Documento66 pagineDevelopment Psychology - Chapter 4 (Santrock)Shaine C.Nessuna valutazione finora
- Urinary System: Excrete Produced Processes Filtration Reabsorption Secretion Excretion ConcentrateDocumento2 pagineUrinary System: Excrete Produced Processes Filtration Reabsorption Secretion Excretion ConcentrateEurasia BlossomNessuna valutazione finora
- Biochemistry (GREEN)Documento18 pagineBiochemistry (GREEN)BRYAN BALDOMERONessuna valutazione finora
- 1) Pathology: Myasthenia Gravis Is An Autoimmune Disease Associated With Antibodies Directed To TheDocumento49 pagine1) Pathology: Myasthenia Gravis Is An Autoimmune Disease Associated With Antibodies Directed To TheAhmad HassanNessuna valutazione finora
- Bahagian A (20 Soalan) : 1. Study The Information BelowDocumento15 pagineBahagian A (20 Soalan) : 1. Study The Information BelowDesmond ChicharitoNessuna valutazione finora
- Myofascial Pain SyndromeDocumento61 pagineMyofascial Pain Syndromeyosra adam100% (1)
- FitokimDocumento16 pagineFitokimAdilah AthtahirahNessuna valutazione finora
- Enzyme Action: Learning Objective: To Describe Different Models ForDocumento28 pagineEnzyme Action: Learning Objective: To Describe Different Models ForMatt BarhamNessuna valutazione finora