Potrebbero piacerti anche
- Healing Crystals For BeginnersDocumento112 pagineHealing Crystals For BeginnersBibliophilia790% (21)
- 5 Kitchen Spices Ebook by Kami McbrideDocumento11 pagine5 Kitchen Spices Ebook by Kami Mcbridenicolas0danek100% (2)
- Anatomy and Physiology II Course OutcomesDocumento50 pagineAnatomy and Physiology II Course OutcomesShirley LourdesNessuna valutazione finora
- Cardiac Cycle 1Documento45 pagineCardiac Cycle 1Ashhal KNessuna valutazione finora
- Lymph SystemDocumento7 pagineLymph SystemghaiathNessuna valutazione finora
- The Kidney: Hemanth P Dialysis Tutor B N Patel Institute of Paramedical & SciencesDocumento46 pagineThe Kidney: Hemanth P Dialysis Tutor B N Patel Institute of Paramedical & SciencesHemanth PrakashNessuna valutazione finora
- Ovarian Cycle & Menestrual CycleDocumento16 pagineOvarian Cycle & Menestrual CycleRolando Ian Maranan RNNessuna valutazione finora
- Respiratory SystemDocumento36 pagineRespiratory SystemPritha Bhuwapaksophon100% (1)
- Causes of Erectile Dysfunction As Explained in AyurvedaDocumento25 pagineCauses of Erectile Dysfunction As Explained in Ayurvedacharanmann9165Nessuna valutazione finora
- Hip JointDocumento25 pagineHip JointHashim OmarNessuna valutazione finora
- Cardiovascular System: More Than Just The HeartDocumento34 pagineCardiovascular System: More Than Just The HearteliseudesafateNessuna valutazione finora
- SPIROMETRYDocumento5 pagineSPIROMETRYJayarubini JeyapalNessuna valutazione finora
- Circulatory System - Part 2 4-8-14 For BBDocumento23 pagineCirculatory System - Part 2 4-8-14 For BBroman7dbNessuna valutazione finora
- M - 126 Structure of Nephron and Function of The Kidney PDFDocumento4 pagineM - 126 Structure of Nephron and Function of The Kidney PDFDr. Tapan Kr. Dutta100% (1)
- Primary Venous Drainage of the BladderDocumento121 paginePrimary Venous Drainage of the BladderTowhid HasanNessuna valutazione finora
- Human Anatomy,: First Edition Mckinley & O'LoughlinDocumento40 pagineHuman Anatomy,: First Edition Mckinley & O'LoughlinAgung PurmanaNessuna valutazione finora
- Blood Cells, Immunity and Blood ClottingDocumento65 pagineBlood Cells, Immunity and Blood Clottingmunaamuummee100% (1)
- Operational K9 TECC Challenges White Paper Final Dated 29 Sep 2015Documento23 pagineOperational K9 TECC Challenges White Paper Final Dated 29 Sep 2015Maikol FloresNessuna valutazione finora
- Oogenesis and Ovarian Cycle, Uterus, Menstrual Cycle ABORTION TERATOLOGYDocumento72 pagineOogenesis and Ovarian Cycle, Uterus, Menstrual Cycle ABORTION TERATOLOGYElizabeth OmobolanleNessuna valutazione finora
- Dr. Aaijaz Ahmed Khan Sr. Lecturer in Anatomy PPSP, UsmDocumento38 pagineDr. Aaijaz Ahmed Khan Sr. Lecturer in Anatomy PPSP, UsmeliseudesafateNessuna valutazione finora
- Membrane Transport Mechanisms and HomeostasisDocumento29 pagineMembrane Transport Mechanisms and Homeostasisramadan100% (1)
- Hyperosmolar Non Ketotic ComaDocumento23 pagineHyperosmolar Non Ketotic Comaprakash gunaNessuna valutazione finora
- I.V Fluid: PathologyDocumento7 pagineI.V Fluid: PathologyruchikaNessuna valutazione finora
- Mechanism of Urine FormationDocumento5 pagineMechanism of Urine FormationovacuteNessuna valutazione finora
- Composition and Function of Blood ComponentsDocumento17 pagineComposition and Function of Blood ComponentsPrakash Kumar Nayak100% (1)
- ID Bug chart-DKDocumento92 pagineID Bug chart-DKNeil M D'SouzaNessuna valutazione finora
- Care For Hemodialysis Patients: Pre, During, Post Dialysis CareDocumento18 pagineCare For Hemodialysis Patients: Pre, During, Post Dialysis CareHemanth PrakashNessuna valutazione finora
- Chick NeurulationDocumento14 pagineChick NeurulationSophia RubiaNessuna valutazione finora
- Pets and PeopleDocumento329 paginePets and PeopleMénades Editorial100% (1)
- The Human Life Begins at ConceptionDocumento12 pagineThe Human Life Begins at ConceptionDavid KrogerNessuna valutazione finora
- Comparing band math equations for measuring suspended material in coastal watersDocumento6 pagineComparing band math equations for measuring suspended material in coastal waterslabibmalikalmhadiNessuna valutazione finora
- Muscle PhysiologyDocumento24 pagineMuscle PhysiologyJorem Paulo LabaoNessuna valutazione finora
- Anatomy and PhysiologyDocumento17 pagineAnatomy and PhysiologychardiginNessuna valutazione finora
- Cell Diffusion & Permeability - Teacher VersionDocumento9 pagineCell Diffusion & Permeability - Teacher VersionTeachLABScINessuna valutazione finora
- Water, Electrolyte and PH BalanceDocumento6 pagineWater, Electrolyte and PH BalancePitambar PoudelNessuna valutazione finora
- Fluid Therapy in Various ConditionsDocumento62 pagineFluid Therapy in Various ConditionsAndy F MonroeNessuna valutazione finora
- Urine Formation Steps Filtration: Dr. Sumaiya KhanDocumento4 pagineUrine Formation Steps Filtration: Dr. Sumaiya KhanZeleen Ann SaberonNessuna valutazione finora
- Threshold and RecruitmentDocumento16 pagineThreshold and RecruitmentShamaine Anne SalvadorNessuna valutazione finora
- Structure and Functions of The Major Types of Blood VesselsDocumento6 pagineStructure and Functions of The Major Types of Blood VesselsCharls DeimoyNessuna valutazione finora
- Chapter 26: The Urinary System: Chapter Objectives Overview of Kidney FunctionDocumento14 pagineChapter 26: The Urinary System: Chapter Objectives Overview of Kidney FunctionAnkit SwainNessuna valutazione finora
- 16/09/2015 1 Mujtaba AshrafDocumento44 pagine16/09/2015 1 Mujtaba Ashrafதீரன் சக்திவேல்Nessuna valutazione finora
- Cell StructureDocumento51 pagineCell Structureapi-233187566Nessuna valutazione finora
- 1st Lecture On The Histology of Skin by Dr. RoomiDocumento14 pagine1st Lecture On The Histology of Skin by Dr. RoomiMudassar RoomiNessuna valutazione finora
- Histology of Glands: Dr. Zana TahseenDocumento28 pagineHistology of Glands: Dr. Zana TahseenAhmed JawdetNessuna valutazione finora
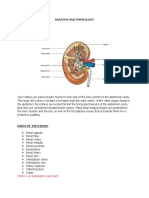
- Anatomy and Physiology - Kidney - ForDocumento3 pagineAnatomy and Physiology - Kidney - ForKristelle ModalesNessuna valutazione finora
- Lymphatic SystemDocumento29 pagineLymphatic SystemlecturioNessuna valutazione finora
- Presented By: DR Sharmila G SDocumento76 paginePresented By: DR Sharmila G SSharmila Shivakumar G SNessuna valutazione finora
- Chapter25 Urinary SystemDocumento9 pagineChapter25 Urinary Systemkikajet23Nessuna valutazione finora
- CV SYSTEM ANATOMYDocumento42 pagineCV SYSTEM ANATOMYJerilee SoCute WattsNessuna valutazione finora
- Anatomy and Physiology MouthDocumento4 pagineAnatomy and Physiology MouthAndrew BimbusNessuna valutazione finora
- Renal Physiology: Tubular ProcessingDocumento6 pagineRenal Physiology: Tubular ProcessingUloko ChristopherNessuna valutazione finora
- Anatomy of Urinary SystemDocumento34 pagineAnatomy of Urinary Systemhana100% (1)
- Digestive System PowerpointDocumento19 pagineDigestive System Powerpointapi-277974939Nessuna valutazione finora
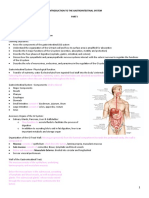
- 1 Introduction To The Gastrointestinal SystemDocumento7 pagine1 Introduction To The Gastrointestinal SystemLinh Phan100% (1)
- Thyroid Gland BasicsDocumento25 pagineThyroid Gland BasicsSreekar DevarakondaNessuna valutazione finora
- Bped 65 Urinary SystemDocumento26 pagineBped 65 Urinary SystemDollie May Maestre-TejidorNessuna valutazione finora
- Macbeth Summery.Documento2 pagineMacbeth Summery.Kafayetsaimum100% (1)
- Blood: Beenish Gul Khattak Lecturer Pharmacy Abasyn University PeshawarDocumento66 pagineBlood: Beenish Gul Khattak Lecturer Pharmacy Abasyn University PeshawarEsperanza KtkNessuna valutazione finora
- Pancreatic Juice SecretionDocumento3 paginePancreatic Juice SecretionBilal SaidNessuna valutazione finora
- Embryonic Period: Dr. Khin Ma MADocumento34 pagineEmbryonic Period: Dr. Khin Ma MAIbrahim FoondunNessuna valutazione finora
- Classification of MusclesDocumento50 pagineClassification of MusclesNoor HaleemNessuna valutazione finora
- Properties of Cardiac MuscleDocumento5 pagineProperties of Cardiac Musclekiedd_04100% (4)
- Respiratory SystemDocumento56 pagineRespiratory SystemLorenz PasensyosoNessuna valutazione finora
- 3rd Lecture On Skeletal Muscle Physiology by DR - RoomiDocumento21 pagine3rd Lecture On Skeletal Muscle Physiology by DR - RoomiMudassar Roomi67% (3)
- Blood Vessels Review SheetDocumento6 pagineBlood Vessels Review Sheet9Speed93% (14)
- BSC Nursing - Cardiac OutputDocumento10 pagineBSC Nursing - Cardiac OutputDIKSHA RANI DEKANessuna valutazione finora
- Liver Anatomy and PhysiologyDocumento23 pagineLiver Anatomy and PhysiologyChiragNessuna valutazione finora
- Digestive SystemDocumento36 pagineDigestive Systembjbitscs21Nessuna valutazione finora
- Mechanism and Regulation of Urine Formation Part 1Documento5 pagineMechanism and Regulation of Urine Formation Part 1Zi .aNessuna valutazione finora
- AnemiaDocumento6 pagineAnemiaHemanth PrakashNessuna valutazione finora
- Anti GBM DiseaseDocumento36 pagineAnti GBM DiseaseHemanth PrakashNessuna valutazione finora
- Acute Renal Failure or Acute Kidney InjuryDocumento8 pagineAcute Renal Failure or Acute Kidney InjuryHemanth PrakashNessuna valutazione finora
- Alport'S SyndromeDocumento8 pagineAlport'S SyndromeHemanth PrakashNessuna valutazione finora
- Acute NephritisDocumento6 pagineAcute NephritisHemanth PrakashNessuna valutazione finora
- Dialysis: Hemanth P Author atDocumento8 pagineDialysis: Hemanth P Author atHemanth PrakashNessuna valutazione finora
- Access RecirculationDocumento2 pagineAccess RecirculationHemanth PrakashNessuna valutazione finora
- Acute Renal Failure or Acute Kidney InjuryDocumento3 pagineAcute Renal Failure or Acute Kidney InjuryHemanth PrakashNessuna valutazione finora
- Cardiac Diseases in CKDDocumento4 pagineCardiac Diseases in CKDHemanth PrakashNessuna valutazione finora
- 2.nephrotic SyndromeDocumento7 pagine2.nephrotic SyndromeHemanth PrakashNessuna valutazione finora
- Cardiac Complications in CKDDocumento37 pagineCardiac Complications in CKDHemanth PrakashNessuna valutazione finora
- Ascites in Dialysis Patients 24.8.18Documento10 pagineAscites in Dialysis Patients 24.8.18Hemanth PrakashNessuna valutazione finora
- Cardiovascular Complication in CKD 28.8.18Documento65 pagineCardiovascular Complication in CKD 28.8.18Hemanth PrakashNessuna valutazione finora
- Donor EvaluationDocumento20 pagineDonor EvaluationHemanth PrakashNessuna valutazione finora
- Dialysis Amyloidisis 27.8.18Documento9 pagineDialysis Amyloidisis 27.8.18Hemanth PrakashNessuna valutazione finora
- Continuous Renal Replacement TherapiesDocumento35 pagineContinuous Renal Replacement TherapiesHemanth PrakashNessuna valutazione finora
- Neurological Complications of Uremia and Dialysis ExplainedDocumento55 pagineNeurological Complications of Uremia and Dialysis ExplainedHemanth PrakashNessuna valutazione finora
- MicturitionDocumento6 pagineMicturitionHemanth PrakashNessuna valutazione finora
- Physiology of Bladder FunctionDocumento13 paginePhysiology of Bladder FunctionHemanth PrakashNessuna valutazione finora
- Domestic Goat GuideDocumento66 pagineDomestic Goat Guideascui7434Nessuna valutazione finora
- Ventricular Septal Defect PresentationDocumento12 pagineVentricular Septal Defect Presentationsamchow262100% (1)
- Phospholipid Scramblase An UpdateDocumento7 paginePhospholipid Scramblase An UpdateUmmiNessuna valutazione finora
- CH 11 Heart NotesDocumento2 pagineCH 11 Heart Notesummnicole0% (1)
- BlastulaDocumento4 pagineBlastulaCyanDesNessuna valutazione finora
- Year of The Dog 2018Documento60 pagineYear of The Dog 2018angora6750232Nessuna valutazione finora
- Agriculture SBA - Broiler Production - CDocumento8 pagineAgriculture SBA - Broiler Production - Calex kingNessuna valutazione finora
- Getting Out of ItDocumento36 pagineGetting Out of ItNavneet ChaubeyNessuna valutazione finora
- By Taking On Poliovirus, Marguerite Vogt Transformed The Study of All VirusesDocumento8 pagineBy Taking On Poliovirus, Marguerite Vogt Transformed The Study of All VirusesYoNessuna valutazione finora
- Assignment 3Documento2 pagineAssignment 3Sophie Siti SNessuna valutazione finora
- 158-Article Text-220-1-10-20181218Documento4 pagine158-Article Text-220-1-10-20181218Fania Aulia RahmaNessuna valutazione finora
- The Midlife: Henrietta V. Cabado-Española, MD, FPPADocumento7 pagineThe Midlife: Henrietta V. Cabado-Española, MD, FPPADoc Prince CaballeroNessuna valutazione finora
- Skin and Its Applied AnatomyDocumento56 pagineSkin and Its Applied AnatomyShailesh PatilNessuna valutazione finora
- Anatomy and PhysiologyDocumento5 pagineAnatomy and PhysiologyAica LupenaNessuna valutazione finora
- Kuliah Trauma UG PDFDocumento44 pagineKuliah Trauma UG PDFFazliah FzNessuna valutazione finora
- LECTURE 1 - Respiratory SystemDocumento1 paginaLECTURE 1 - Respiratory SystemJeremiah AndalNessuna valutazione finora
- Animal Research Facility Design and RequirementsDocumento8 pagineAnimal Research Facility Design and RequirementsLuiza Mae CastroNessuna valutazione finora
- Oral RevalidaDocumento98 pagineOral RevalidaJay ArNessuna valutazione finora
- Dental PulpDocumento19 pagineDental Pulpxehd10100% (5)
- AAAP Avian Disease Manual 8th EditionDocumento652 pagineAAAP Avian Disease Manual 8th EditionMarilyn Soto GuillénNessuna valutazione finora
- How To Treat Sick Chickens (Quick Guide) - The Poultry ShopDocumento7 pagineHow To Treat Sick Chickens (Quick Guide) - The Poultry ShopReport PlussNessuna valutazione finora
- Procedures Used in The Management of DystociaDocumento17 pagineProcedures Used in The Management of DystociaVeterinary Online InstituteNessuna valutazione finora
- Bharad Vaja SanaDocumento7 pagineBharad Vaja SanaLomombNessuna valutazione finora