Potrebbero piacerti anche
- Da Antiguidade à Redescoberta das Leis de MendelDa EverandDa Antiguidade à Redescoberta das Leis de MendelNessuna valutazione finora
- Potencialidades Da Moringa Oleifera Lam. No Semiárido Nordestino BrasileiroDa EverandPotencialidades Da Moringa Oleifera Lam. No Semiárido Nordestino BrasileiroNessuna valutazione finora
- Catálise enzimática - mecanismos e exemplosDocumento72 pagineCatálise enzimática - mecanismos e exemplosDayane FaleiroNessuna valutazione finora
- Processo produtivo e comercialização da resina de pinusDocumento22 pagineProcesso produtivo e comercialização da resina de pinusTamires PenidoNessuna valutazione finora
- Curso: Mestrado em Desenvolvimento Humano e Educação: Armando Domingos Cesar Mussa Maia Chacha Banco MondolaDocumento19 pagineCurso: Mestrado em Desenvolvimento Humano e Educação: Armando Domingos Cesar Mussa Maia Chacha Banco MondolaArmando DomingosNessuna valutazione finora
- Adaptações anatômicas das plantas à secaDocumento7 pagineAdaptações anatômicas das plantas à secaLIANDRA BRASIL PIRESNessuna valutazione finora
- Ebook EDUCGEN - Melhoramento Genético de Plantas para Resistência A PragasDocumento129 pagineEbook EDUCGEN - Melhoramento Genético de Plantas para Resistência A PragasClaudius LuciusNessuna valutazione finora
- Metabolismo de Bases NitrogenadasDocumento42 pagineMetabolismo de Bases NitrogenadasBeatriz Dias100% (1)
- Trabalho de MicrobiologiaDocumento35 pagineTrabalho de MicrobiologiaHélen Cristina100% (5)
- Cereais, leguminosas e qualidade alimentarDocumento28 pagineCereais, leguminosas e qualidade alimentarFelo André IdrisseNessuna valutazione finora
- Exercicios de Ecologia Geral e HumanaDocumento15 pagineExercicios de Ecologia Geral e HumanaHermeneGildoAntónioMmiraNessuna valutazione finora
- Efeitos da salinidade nas plantasDocumento18 pagineEfeitos da salinidade nas plantasEllen OliveiraNessuna valutazione finora
- Respiração celular processoDocumento5 pagineRespiração celular processoRosalina Barros100% (1)
- Mangueira: Guia completo sobre a árvore e fruto tropicalDocumento10 pagineMangueira: Guia completo sobre a árvore e fruto tropicalDayane MacedoNessuna valutazione finora
- Fotomorfogênese: pigmentos que absorvem a luz solarDocumento12 pagineFotomorfogênese: pigmentos que absorvem a luz solarantonio ferrao verissimoNessuna valutazione finora
- A Importância de Aulas PráticasDocumento19 pagineA Importância de Aulas Práticasmoacirtomaz2760100% (3)
- Adubação FoliarDocumento8 pagineAdubação FoliarFabíola EstrelaNessuna valutazione finora
- Fotossíntese C3-C4-CAMDocumento13 pagineFotossíntese C3-C4-CAMHelder LucenaNessuna valutazione finora
- Metabolismo CelularDocumento9 pagineMetabolismo CelularCleyton SantosNessuna valutazione finora
- Produção de Feijão NhembaDocumento13 pagineProdução de Feijão NhembaMunir Massarote Jr.Nessuna valutazione finora
- Trabalho Final-Metodologia de ProjetoDocumento9 pagineTrabalho Final-Metodologia de ProjetoTamires ToniotiNessuna valutazione finora
- Coevolução inseto-plantaDocumento8 pagineCoevolução inseto-plantaAna Paula MendesNessuna valutazione finora
- Musaceae - Família da bananaDocumento19 pagineMusaceae - Família da bananaLuke CarricondeNessuna valutazione finora
- (051811220523) Propriedades Organolepticas MadeiraDocumento5 pagine(051811220523) Propriedades Organolepticas MadeiraRodrigo AndradeNessuna valutazione finora
- Efeito de Diferentes Doses de Etileno em BananasDocumento47 pagineEfeito de Diferentes Doses de Etileno em Bananasvaz Francisco albertoNessuna valutazione finora
- Apontamento AgropecuáriaDocumento3 pagineApontamento AgropecuáriaSalimo TidonioNessuna valutazione finora
- Efeitos de Plantas Nativas no Controlo de Pragas e Rendimento de FeijãoDocumento71 pagineEfeitos de Plantas Nativas no Controlo de Pragas e Rendimento de FeijãoDouglas NorbertoNessuna valutazione finora
- HomeostaseDocumento5 pagineHomeostaseTiê FelixNessuna valutazione finora
- Conservação de germoplasma e melhoramento vegetalDocumento87 pagineConservação de germoplasma e melhoramento vegetalForte CellNessuna valutazione finora
- PALOP MozambiqueDocumento94 paginePALOP MozambiqueAlfredo Bosque BondeNessuna valutazione finora
- Educação Física TMBDocumento4 pagineEducação Física TMBJean Freitas LimaNessuna valutazione finora
- Tecnicas de Conservacao de AlimentosDocumento16 pagineTecnicas de Conservacao de AlimentosBarbudo LGNessuna valutazione finora
- Levantamento de Areas ProtegidasDocumento11 pagineLevantamento de Areas ProtegidasIvan AnibalNessuna valutazione finora
- Seleccionar, Preparar e Armazenar SementesDocumento64 pagineSeleccionar, Preparar e Armazenar Sementescondedavisconde100% (1)
- Acessibilidade em escolas e a inclusão de alunos com deficiência motoraDocumento8 pagineAcessibilidade em escolas e a inclusão de alunos com deficiência motoraEdmilson Estevão AzevedoNessuna valutazione finora
- Relatório final de pesquisa sobre alho e cebola em MoçambiqueDocumento27 pagineRelatório final de pesquisa sobre alho e cebola em Moçambiquecalton albanoNessuna valutazione finora
- Os Processos Fisiológicos Da Regeneração e TransplantaçãoDocumento13 pagineOs Processos Fisiológicos Da Regeneração e TransplantaçãoAntónio Pereira100% (1)
- Matéria-Prima - MandiocaDocumento15 pagineMatéria-Prima - MandiocaDANIELLE RAQUEL NASCIMENTO DA SILVANessuna valutazione finora
- Modulo 1 - Introdução A AnatomiaDocumento26 pagineModulo 1 - Introdução A AnatomialeealencarNessuna valutazione finora
- Ensino Matemática Educação Infantil CampoDocumento5 pagineEnsino Matemática Educação Infantil CampoJoão CamposNessuna valutazione finora
- Desafios da Extensão Rural para o Desenvolvimento SustentávelDocumento23 pagineDesafios da Extensão Rural para o Desenvolvimento SustentávelWeverton João VictorNessuna valutazione finora
- A importância da ética na formação do administradorDocumento16 pagineA importância da ética na formação do administradorEva Ferreira DuarteNessuna valutazione finora
- Aula 4. Mecanismos de Fixação de CO2 em PlantasDocumento7 pagineAula 4. Mecanismos de Fixação de CO2 em PlantasFrancisco Rosario JaimeNessuna valutazione finora
- Biologia Das Algas e Plantas PDFDocumento49 pagineBiologia Das Algas e Plantas PDFFCiências0% (1)
- Tratamento de SementesDocumento28 pagineTratamento de SementesAtarissis Dias100% (1)
- Lipídios: Extração e TeorDocumento17 pagineLipídios: Extração e TeorLeonardo Antunes SodreNessuna valutazione finora
- Graficos Estatisticos JayDocumento13 pagineGraficos Estatisticos JayEma Nuel100% (1)
- Parcelas PerdidasDocumento7 pagineParcelas PerdidasThalita Do Bem67% (3)
- Higiene e Tecnologia de Conservação de PescadoDocumento24 pagineHigiene e Tecnologia de Conservação de PescadoFranklin Da Joana MuceloNessuna valutazione finora
- Frutas e citrinos na 10a classeDocumento7 pagineFrutas e citrinos na 10a classeHelder CoelhoNessuna valutazione finora
- Didática e Práticas de Ensino e A Abordagem Da Diversidade Sociocultural Na Escola PDFDocumento21 pagineDidática e Práticas de Ensino e A Abordagem Da Diversidade Sociocultural Na Escola PDFSyndhia Rodrigues100% (1)
- CarbohidratosDocumento12 pagineCarbohidratosabdul lingeNessuna valutazione finora
- Horta Escolar: Uma estratégia de ecoalfabetizaçãoDocumento11 pagineHorta Escolar: Uma estratégia de ecoalfabetizaçãoElayne SantosNessuna valutazione finora
- Os principais grupos de artrópodes: características, classificação e importânciaDocumento13 pagineOs principais grupos de artrópodes: características, classificação e importânciaMunlhena ArturNessuna valutazione finora
- Definição o e Importância Da Silvicultura FlorestalDocumento34 pagineDefinição o e Importância Da Silvicultura FlorestalMarcos AlbertoNessuna valutazione finora
- Intemperismo e PedogêneseDocumento24 pagineIntemperismo e Pedogêneseandressaoliveira2301Nessuna valutazione finora
- Trabalho Colheita e Pós Colheita de MilhoDocumento22 pagineTrabalho Colheita e Pós Colheita de MilhoFagner Fernandes da Silva67% (3)
- Atributos Físicos, Químicos e Biológicos Do SoloDocumento22 pagineAtributos Físicos, Químicos e Biológicos Do SoloSamuel SantosNessuna valutazione finora
- Pesquisas e práticas de ensino de um professor biólogo em formaçãoDa EverandPesquisas e práticas de ensino de um professor biólogo em formaçãoNessuna valutazione finora
- A Temática Ambiental na Escola e os Artefatos da Indústria CulturalDa EverandA Temática Ambiental na Escola e os Artefatos da Indústria CulturalNessuna valutazione finora
- Regulamento sobre Qualidade Água Consumo HumanoDocumento14 pagineRegulamento sobre Qualidade Água Consumo HumanoAngelo ThiagoNessuna valutazione finora
- O debate sobre o salário mínimo em MoçambiqueDocumento17 pagineO debate sobre o salário mínimo em MoçambiqueAngelo Thiago100% (2)
- BioetanolDocumento316 pagineBioetanolKadu MeloNessuna valutazione finora
- 5.1. Difusao Interna. ExerciciosDocumento13 pagine5.1. Difusao Interna. ExerciciosAngelo ThiagoNessuna valutazione finora
- Titulação ConductométricaDocumento9 pagineTitulação ConductométricaAngelo ThiagoNessuna valutazione finora
- Conversores A-D e D-ADocumento21 pagineConversores A-D e D-AAngelo ThiagoNessuna valutazione finora
- 06 MaximaTransferenciadePotenciaDocumento6 pagine06 MaximaTransferenciadePotenciaNelson Knak NetoNessuna valutazione finora
- Química 10 ClasseDocumento110 pagineQuímica 10 ClasseAngelo Thiago100% (8)
- Trabalho Tecnicas Empesariais IDocumento12 pagineTrabalho Tecnicas Empesariais IAngelo ThiagoNessuna valutazione finora
- 1 - Transistores II - v1.0Documento32 pagine1 - Transistores II - v1.0Wilson Tadeu AssisNessuna valutazione finora
- Adsorcao 2016 - Parte 1.1Documento8 pagineAdsorcao 2016 - Parte 1.1Angelo ThiagoNessuna valutazione finora
- Atividades de Alongamento CorporalDocumento32 pagineAtividades de Alongamento CorporalAngelo ThiagoNessuna valutazione finora
- Processo de Fabricação de CimentoDocumento18 pagineProcesso de Fabricação de CimentoMarcia G. SilvaNessuna valutazione finora
- PROd CimentoDocumento8 paginePROd CimentoAngelo ThiagoNessuna valutazione finora
- Mecanismos de Reações Orgânicas - Marcelo Pelisson PDFDocumento92 pagineMecanismos de Reações Orgânicas - Marcelo Pelisson PDFSoraya Bio VetNessuna valutazione finora
- TQ - Calores EspecificosDocumento20 pagineTQ - Calores EspecificosAngelo ThiagoNessuna valutazione finora
- Capitulo3 Custos de ManutencaoDocumento32 pagineCapitulo3 Custos de ManutencaoAngelo ThiagoNessuna valutazione finora
- Determinação da viscosidade de óleo usado e não usadoDocumento9 pagineDeterminação da viscosidade de óleo usado e não usadoAngelo ThiagoNessuna valutazione finora
- Análise dimensional e similaridade em fenômenos de transporteDocumento8 pagineAnálise dimensional e similaridade em fenômenos de transporteadolfoeiras6714Nessuna valutazione finora
- Formas de Transferência de Calor e Equações de ConduçãoDocumento7 pagineFormas de Transferência de Calor e Equações de ConduçãoAngelo ThiagoNessuna valutazione finora
- Produção Do AçoDocumento20 pagineProdução Do AçoPedro Polastri PatriotaNessuna valutazione finora
- Acido NitricoDocumento12 pagineAcido NitricoAngelo ThiagoNessuna valutazione finora
- Cap6 AlgoritmosOrdenacaoDocumento9 pagineCap6 AlgoritmosOrdenacaoAngelo ThiagoNessuna valutazione finora
- Laboratório Química Dos Elementos RepresentativosDocumento8 pagineLaboratório Química Dos Elementos RepresentativosAngelo ThiagoNessuna valutazione finora
- Produção Do AçoDocumento20 pagineProdução Do AçoPedro Polastri PatriotaNessuna valutazione finora
- Pendulo e Modulo de TorsaoDocumento10 paginePendulo e Modulo de TorsaoAngelo ThiagoNessuna valutazione finora
- Port Aula 01Documento86 paginePort Aula 01victooriafNessuna valutazione finora
- Mecanismos de SplicingDocumento1 paginaMecanismos de SplicingusrjpfptNessuna valutazione finora
- MECÂNICA DOS SOLOS I - LISTA DE EXERCÍCIOSDocumento1 paginaMECÂNICA DOS SOLOS I - LISTA DE EXERCÍCIOSLeonardo Santa RitaNessuna valutazione finora
- A autoestima: desenvolver a confiança em siDocumento26 pagineA autoestima: desenvolver a confiança em siTeisa Rosa100% (17)
- 3 Banhos para Ficar Atraente Homens Rastejando em 24 Horas!Documento1 pagina3 Banhos para Ficar Atraente Homens Rastejando em 24 Horas!Catarina MateusNessuna valutazione finora
- Técnicas de Aramação do BonsaiDocumento5 pagineTécnicas de Aramação do Bonsaifegon3001Nessuna valutazione finora
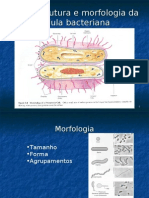
- Ultraestrutura e Morfologia Bacteriana 4Documento28 pagineUltraestrutura e Morfologia Bacteriana 4MaxD2099100% (1)
- Primeira TopicaDocumento5 paginePrimeira TopicaGustavo Moreira AlvesNessuna valutazione finora
- Avaliação de Geografia 2º ANODocumento4 pagineAvaliação de Geografia 2º ANOFRANCILENE84% (115)
- Avaliação Do Estado Do Conhecimento Da Biodiversidade BrasileiraDocumento271 pagineAvaliação Do Estado Do Conhecimento Da Biodiversidade Brasileiramonteirorv9740Nessuna valutazione finora
- Modelos Geocêntrico e HeliocêntricoDocumento19 pagineModelos Geocêntrico e HeliocêntricoMárcia Beatriz Cruz Sousa100% (1)
- Investigação geológica em projetos de engenhariaDocumento2 pagineInvestigação geológica em projetos de engenhariaKaren LeopoldinoNessuna valutazione finora
- Topiramato na profilaxia de enxaqueca em crianças: revisão sistemáticaDocumento31 pagineTopiramato na profilaxia de enxaqueca em crianças: revisão sistemáticaMileni E MarcusNessuna valutazione finora
- Evolução da atmosfera terrestre e surgimento da vidaDocumento4 pagineEvolução da atmosfera terrestre e surgimento da vidaMaria Ol100% (1)
- Classificação de seres vivos ao longo da históriaDocumento5 pagineClassificação de seres vivos ao longo da históriaMargarida PassanhaNessuna valutazione finora
- Determinação da posição geográfica através de coordenadasDocumento221 pagineDeterminação da posição geográfica através de coordenadasJunior CarvalhoNessuna valutazione finora
- 10 Coisas Que As Pessoas Muito Intuitivas Fazem DiferenteDocumento4 pagine10 Coisas Que As Pessoas Muito Intuitivas Fazem DiferenteVanessa SilvaNessuna valutazione finora
- Planificação História e Geografia 5º Ano - Século XV e XVIDocumento3 paginePlanificação História e Geografia 5º Ano - Século XV e XVIramiromarquesNessuna valutazione finora
- Atividade Avaliativa de CiênciasDocumento5 pagineAtividade Avaliativa de CiênciasDevanir YoneharaNessuna valutazione finora
- Checklist Do Pará - EspéciesDocumento45 pagineChecklist Do Pará - EspéciesCardosoNessuna valutazione finora
- Importância e conceitos sobre forrageiras tropicaisDocumento33 pagineImportância e conceitos sobre forrageiras tropicaisRubens JuniorNessuna valutazione finora
- WINGMAKERS - TraduçãoDocumento51 pagineWINGMAKERS - TraduçãoJosé Carlos Silva Costa100% (1)
- O Clima Na Península IbéricaDocumento3 pagineO Clima Na Península IbéricaSónia RibeiroNessuna valutazione finora
- DO Abilio Coppede CorrigidaDocumento202 pagineDO Abilio Coppede CorrigidaodontosidyNessuna valutazione finora
- A Origem Do Sistema SolarDocumento3 pagineA Origem Do Sistema SolaranaisabelfernandesNessuna valutazione finora
- Comportamentalismo - ResumoDocumento9 pagineComportamentalismo - ResumoCláudiaDaSilvaSoaresFeitosaNessuna valutazione finora
- Lista 9 Pob.Documento4 pagineLista 9 Pob.Carllos LimmaNessuna valutazione finora
- Estaca Tipo FRANKIDocumento42 pagineEstaca Tipo FRANKIJamilson Oliveira50% (2)
- COGNIÇÃO SOCIAL E PSICOPATIADocumento10 pagineCOGNIÇÃO SOCIAL E PSICOPATIAandreashellfireNessuna valutazione finora
- Colisões e conservação da quantidade de movimentoDocumento2 pagineColisões e conservação da quantidade de movimentoCarlos Jorge MedeirosNessuna valutazione finora