Potrebbero piacerti anche
- Devlin Biochemistry With Clinical CorrelationsDocumento1.241 pagineDevlin Biochemistry With Clinical CorrelationsJorge100% (5)
- 7Documento15 pagine7AmaniNessuna valutazione finora
- CryopreservationDocumento47 pagineCryopreservationanitha pathlavath100% (1)
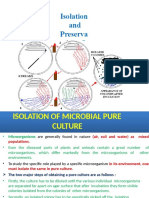
- Lecture 8-Isolation and Enrichment TechniquesDocumento25 pagineLecture 8-Isolation and Enrichment TechniquesSELVI A100% (3)
- Stages of BioprocessingDocumento8 pagineStages of BioprocessingMd Shahjalal Sagor0% (1)
- Personalized Package Attunement List WWW - Livingreikiacademy.co - UkDocumento47 paginePersonalized Package Attunement List WWW - Livingreikiacademy.co - Ukriddhisnewworld67% (6)
- S-BIOL327 INTRODUCTORY GENETICS LECTURE MEB23 (2nd Semester SY 2020-2021) Enabling Assessment - Gene RegulationDocumento3 pagineS-BIOL327 INTRODUCTORY GENETICS LECTURE MEB23 (2nd Semester SY 2020-2021) Enabling Assessment - Gene RegulationGenghis SupnetNessuna valutazione finora
- Chapter 28 Regulation of Gene Expression: Multiple Choice QuestionsDocumento11 pagineChapter 28 Regulation of Gene Expression: Multiple Choice QuestionsKhristian Dom Ruam Cadabona40% (5)
- Strain ImprovementDocumento122 pagineStrain ImprovementRishanam RishanaNessuna valutazione finora
- Strain ImprovementDocumento122 pagineStrain ImprovementKhushi GuptaNessuna valutazione finora
- WINSEM2019-20 BIT1007 ETH VL2019205000968 Reference Material I 08-Jan-2020 PreservationDocumento41 pagineWINSEM2019-20 BIT1007 ETH VL2019205000968 Reference Material I 08-Jan-2020 PreservationManish GiriNessuna valutazione finora
- Isolation Screening PDFDocumento23 pagineIsolation Screening PDFMass ManiNessuna valutazione finora
- 2019 BT 204-Module 1 (PartD)Documento32 pagine2019 BT 204-Module 1 (PartD)Leon IttiachenNessuna valutazione finora
- Preservation of Microorganisms 8Documento33 paginePreservation of Microorganisms 8lalithaNessuna valutazione finora
- Strain ImprovementDocumento40 pagineStrain ImprovementSiti khaizatul minnah100% (1)
- Isolationandpreservation of MoDocumento31 pagineIsolationandpreservation of MopraneethasruthiNessuna valutazione finora
- Algal BiomassDocumento25 pagineAlgal Biomassvicky vigneshNessuna valutazione finora
- Preservationof Industrially Important MicroorganismsDocumento18 paginePreservationof Industrially Important Microorganismsjeevalakshmanan29Nessuna valutazione finora
- Culture Preservation TechniquesDocumento16 pagineCulture Preservation TechniquesDr. Kalavati Prajapati50% (2)
- Embryo Rescue, Somaclonal Variation, CryopreservationDocumento32 pagineEmbryo Rescue, Somaclonal Variation, CryopreservationTabada NickyNessuna valutazione finora
- Probiotics: Dr. Arun SIIP Fellow Sctimst-TimedDocumento16 pagineProbiotics: Dr. Arun SIIP Fellow Sctimst-TimedArunNessuna valutazione finora
- Recycling 3Documento29 pagineRecycling 3adhamelthnNessuna valutazione finora
- Stock Culture and Its MaintenanceDocumento4 pagineStock Culture and Its MaintenanceSuresh Mg100% (2)
- Culture Media and Isolation Method ActivityDocumento3 pagineCulture Media and Isolation Method ActivityJake Sim100% (1)
- Pure Culture TechniquesDocumento33 paginePure Culture TechniquesRamachari K VNessuna valutazione finora
- Food PreservationDocumento72 pagineFood PreservationEkta KasanaNessuna valutazione finora
- Preservation of Microbial CultureDocumento4 paginePreservation of Microbial CulturePoojaswini ReddyNessuna valutazione finora
- Lecture 3Documento20 pagineLecture 3Biotechnology IIUM Kuantan100% (5)
- Unit Operation & Unit Process in BiotechnologyDocumento52 pagineUnit Operation & Unit Process in BiotechnologyKailash Kumar100% (1)
- Advanced Bioprocess Engineering: Lecturer Dr. Kamal E. M. Elkahlout Assistant Prof. of BiotechnologyDocumento54 pagineAdvanced Bioprocess Engineering: Lecturer Dr. Kamal E. M. Elkahlout Assistant Prof. of Biotechnologybelete fentayeNessuna valutazione finora
- Pure Culture, Preservation and Maintance Practical.Documento17 paginePure Culture, Preservation and Maintance Practical.Saim Sabeeh0% (1)
- Microbial Protein Production/ Single Cell Protein: - Class TeacherDocumento25 pagineMicrobial Protein Production/ Single Cell Protein: - Class TeacherFaiza NoorNessuna valutazione finora
- Production of VaccinesDocumento95 pagineProduction of Vaccinessahana shanbhagNessuna valutazione finora
- AntibioticsDocumento20 pagineAntibioticsFazal AminNessuna valutazione finora
- Inoculum BestDocumento6 pagineInoculum BestSabarishNessuna valutazione finora
- Fermentation TechnologyDocumento14 pagineFermentation TechnologyUmarNessuna valutazione finora
- Biopharmaceutical 1Documento78 pagineBiopharmaceutical 1Pharmacy Admission ExpertNessuna valutazione finora
- Isolation & Preservation Methods For Pure Culture: Deepankar Ratha, Assistant Professor, CUTM, RayagadaDocumento34 pagineIsolation & Preservation Methods For Pure Culture: Deepankar Ratha, Assistant Professor, CUTM, RayagadaArman KhanxNessuna valutazione finora
- Bioprocess Technology 2022Documento47 pagineBioprocess Technology 2022Sowjanya NekuriNessuna valutazione finora
- Pharmaceutical Microbiology MBO-133 3 CreditDocumento39 paginePharmaceutical Microbiology MBO-133 3 CreditSmk Mahmud KoliNessuna valutazione finora
- Fermentation: Cruz, Tiffany Era, Emil D. Humarang, JelainDocumento24 pagineFermentation: Cruz, Tiffany Era, Emil D. Humarang, JelainTimothy JonesNessuna valutazione finora
- MSC Ind. Biotech. Syllabus (2008-09)Documento4 pagineMSC Ind. Biotech. Syllabus (2008-09)Dr-Alok Prasad DasNessuna valutazione finora
- Conservation of Embryo and Ovules 1Documento41 pagineConservation of Embryo and Ovules 1INDRA RACHMAWATINessuna valutazione finora
- Industrial MCB (DR Etim)Documento35 pagineIndustrial MCB (DR Etim)NAJAATU SHEHU HADINessuna valutazione finora
- Down Stream ProcessingDocumento44 pagineDown Stream ProcessingShifa ChoudharyNessuna valutazione finora
- Maintenance and Preservation of Microbial CulturesDocumento5 pagineMaintenance and Preservation of Microbial CulturesSumit KumarNessuna valutazione finora
- Unit 3Documento39 pagineUnit 3Prabhmeet GroverNessuna valutazione finora
- Week 9Documento15 pagineWeek 9Fernanda QuelNessuna valutazione finora
- Marin Maged 187Documento12 pagineMarin Maged 187Marin MagedNessuna valutazione finora
- What Is Bioprocess?Documento28 pagineWhat Is Bioprocess?sanyukta sinhaNessuna valutazione finora
- "Strategies Used To" Optimize Plant Product Yield: Culture ConditionsDocumento10 pagine"Strategies Used To" Optimize Plant Product Yield: Culture Conditionssaahiba16Nessuna valutazione finora
- Vaccine ProductionDocumento63 pagineVaccine ProductionFaduwa AliNessuna valutazione finora
- Upstream and DownstDocumento21 pagineUpstream and Downstbiotech181985Nessuna valutazione finora
- InBt121 Module 2Documento127 pagineInBt121 Module 2Dylene EstremosNessuna valutazione finora
- Singlecellprotein 120507100927 Phpapp01Documento31 pagineSinglecellprotein 120507100927 Phpapp01GSNessuna valutazione finora
- Single Ceel Protein Very Good FinalDocumento24 pagineSingle Ceel Protein Very Good FinalNavpreet KaurNessuna valutazione finora
- ANTIBIOTICS PenicillinDocumento17 pagineANTIBIOTICS PenicillinDitto V DavisNessuna valutazione finora
- Mass Production of MicroalgaeDocumento27 pagineMass Production of Microalgaerajiv pathakNessuna valutazione finora
- Plant Tissue CultureDocumento38 paginePlant Tissue CultureayeshasattarsandhuNessuna valutazione finora
- Broch Microbiological TestingDocumento28 pagineBroch Microbiological TestingMudassir Ali QazalbashNessuna valutazione finora
- Kimia Pangan - Fermentasi Dan BioreaktorDocumento53 pagineKimia Pangan - Fermentasi Dan BioreaktorHartati SNessuna valutazione finora
- Activity and Purity of StarterDocumento22 pagineActivity and Purity of StarterRohit PandeyNessuna valutazione finora
- DownstreamDocumento67 pagineDownstreamjoyNessuna valutazione finora
- Canned Citrus Processing: Techniques, Equipment, and Food SafetyDa EverandCanned Citrus Processing: Techniques, Equipment, and Food SafetyNessuna valutazione finora
- SUBJECT: Plant Ecology-I Semester: VI, BS Botany Teacher Name: Dr. Khalid Rasheed Khan Department of Botany Government College MansehraDocumento8 pagineSUBJECT: Plant Ecology-I Semester: VI, BS Botany Teacher Name: Dr. Khalid Rasheed Khan Department of Botany Government College MansehrariddhisnewworldNessuna valutazione finora
- Soil Texture and Soil StructureDocumento15 pagineSoil Texture and Soil StructureriddhisnewworldNessuna valutazione finora
- Soil - Structure Interaction: - Thurumella JayanthDocumento17 pagineSoil - Structure Interaction: - Thurumella JayanthriddhisnewworldNessuna valutazione finora
- Different Types of Soil in India and Their Specific Crops: Made ByDocumento20 pagineDifferent Types of Soil in India and Their Specific Crops: Made ByriddhisnewworldNessuna valutazione finora
- Industrial MicroorganismsDocumento7 pagineIndustrial MicroorganismsriddhisnewworldNessuna valutazione finora
- Advantage of Batch and Continous CultureDocumento3 pagineAdvantage of Batch and Continous CultureriddhisnewworldNessuna valutazione finora
- Gene Regulation - Lac Operon & TRP OperonDocumento25 pagineGene Regulation - Lac Operon & TRP OperonSalam Pradeep Singh75% (12)
- Bacteriology Exam IDocumento7 pagineBacteriology Exam ITorillo KimNessuna valutazione finora
- MSC MicrobiologyDocumento13 pagineMSC MicrobiologyShravani SalunkheNessuna valutazione finora
- Concepts of Genetics Books A La Carte Edition 11th Edition Klug Test BankDocumento8 pagineConcepts of Genetics Books A La Carte Edition 11th Edition Klug Test Banklatifahhanhga7100% (30)
- MCQ in BioDocumento7 pagineMCQ in BioGiovanni Roccaforte100% (1)
- Gene BiotecDocumento6 pagineGene BioteclemtechNessuna valutazione finora
- ICMR JRF 2015 Previous - Model Question Paper - EasybiologyclassDocumento10 pagineICMR JRF 2015 Previous - Model Question Paper - EasybiologyclassSharad GajuryalNessuna valutazione finora
- Molecular Cell Biology Practice QuestionsDocumento9 pagineMolecular Cell Biology Practice Questionssahana2791100% (1)
- Prokaryotic and Eukaryotic Gene RegulationDocumento35 pagineProkaryotic and Eukaryotic Gene RegulationDrew TaryeeNessuna valutazione finora
- Regulación Por AtenuaciónDocumento5 pagineRegulación Por AtenuaciónKaryme Aylín Martinez OliverosNessuna valutazione finora
- Gene RegulationDocumento261 pagineGene RegulationPavithra ShanmugamNessuna valutazione finora
- Biochemistry 7th Edition Campbell Test Bank DownloadDocumento33 pagineBiochemistry 7th Edition Campbell Test Bank Downloaddaddockstudderyxeq100% (34)
- 09 Regulation of Gene ExpressionDocumento42 pagine09 Regulation of Gene ExpressionFirst Anonym877Nessuna valutazione finora
- 424 MicrobiologyDocumento72 pagine424 MicrobiologyVignesh ReddyNessuna valutazione finora
- Preclass Quiz 7 - Fa19 - MOLECULAR BIOLOGY (47940) PDFDocumento3 paginePreclass Quiz 7 - Fa19 - MOLECULAR BIOLOGY (47940) PDFElizabeth DouglasNessuna valutazione finora
- Gene Regulation - Operon Theory - Microbiology - OpenStaxDocumento12 pagineGene Regulation - Operon Theory - Microbiology - OpenStaxAleksandra Sanja MartinovicNessuna valutazione finora
- 07 Control of Gene Expression in ProkaryotesDocumento17 pagine07 Control of Gene Expression in Prokaryotesmustafa aliNessuna valutazione finora
- Histidine Operon LeaderDocumento2 pagineHistidine Operon LeaderVijayant Kumar PandeyNessuna valutazione finora
- Kahoot MCQ - StudentDocumento12 pagineKahoot MCQ - Studentnewyork0218Nessuna valutazione finora
- Cambridge International AS & A Level: Biology October/November 2022 2 HoursDocumento28 pagineCambridge International AS & A Level: Biology October/November 2022 2 HourshudaNessuna valutazione finora
- TRP Operon PresentationDocumento27 pagineTRP Operon PresentationPradeep SinghNessuna valutazione finora
- Biomol BiomedikFera Ibrahim08032018Documento95 pagineBiomol BiomedikFera Ibrahim08032018PMIB Matrikulasi FKUI 2018/2019Nessuna valutazione finora
- Gene Regulation - 2018 PDFDocumento95 pagineGene Regulation - 2018 PDFArhanaNessuna valutazione finora
- Gene Regulation in Prokaryotes 11022017Documento55 pagineGene Regulation in Prokaryotes 11022017Nur Syamimi0% (1)
- TRP OperonDocumento80 pagineTRP OperonIsmi SitiNessuna valutazione finora