Potrebbero piacerti anche
- Desnaturalización de Las ProteinasDocumento13 pagineDesnaturalización de Las ProteinasjaimeNessuna valutazione finora
- La Actina PDFDocumento5 pagineLa Actina PDFItzaetlSantanaM0% (1)
- ENZIMASDocumento2 pagineENZIMASAngel Andres Ayquipa AlvitesNessuna valutazione finora
- Glúcidos y Lípidos PDFDocumento33 pagineGlúcidos y Lípidos PDFLucy MamaniNessuna valutazione finora
- Teoria Macro y MicronutrientesDocumento14 pagineTeoria Macro y MicronutrientesGuadeloupe PalNessuna valutazione finora
- Ex. Sustitutorio de HematologíaDocumento13 pagineEx. Sustitutorio de HematologíaRenato Ferré VelásquezNessuna valutazione finora
- Banco Final HematoDocumento10 pagineBanco Final HematoDaniel BracamonteNessuna valutazione finora
- Examen Hemato IIDocumento3 pagineExamen Hemato IIRenato Ferré VelásquezNessuna valutazione finora
- Caso Clinico N°2-Grupo3aDocumento16 pagineCaso Clinico N°2-Grupo3aRenato Ferré VelásquezNessuna valutazione finora
- Caso Clinico n2 DiagnosticoDocumento16 pagineCaso Clinico n2 DiagnosticoRenato Ferré VelásquezNessuna valutazione finora
- Plantilla de Lámina Histológica 1Documento1 paginaPlantilla de Lámina Histológica 1Renato Ferré VelásquezNessuna valutazione finora
- Atencion Primaria de Salud 2010Documento25 pagineAtencion Primaria de Salud 2010fmh2009Nessuna valutazione finora
- Cuestionario Nº16-HistologiaDocumento1 paginaCuestionario Nº16-HistologiaRenato Ferré VelásquezNessuna valutazione finora
- Funcion de Los LípidosDocumento28 pagineFuncion de Los Lípidos•Estella Bemón•Nessuna valutazione finora
- Velocidad de Reaccion Enzimatica 2023aDocumento20 pagineVelocidad de Reaccion Enzimatica 2023aHUERTA ESPINOZA LESSLYNessuna valutazione finora
- Clases de HormonasDocumento8 pagineClases de HormonasMaría Dalia GonzálezNessuna valutazione finora
- TAMY1Documento1 paginaTAMY1ElvilchesNessuna valutazione finora
- Iniciación/nucleación. en Condiciones de Inanición, El Aumento en La RelaciónDocumento3 pagineIniciación/nucleación. en Condiciones de Inanición, El Aumento en La RelaciónDIEGO CRISTHOPER MALPARTIDA ESPINOZANessuna valutazione finora
- Guia Metabolismo y Enzimas Cuarto Medio 2014Documento6 pagineGuia Metabolismo y Enzimas Cuarto Medio 2014Humberto C. BustosNessuna valutazione finora
- Mapa Conceptual Sindrome MetabolicoDocumento8 pagineMapa Conceptual Sindrome MetabolicoLeydi Portalanza VelardeNessuna valutazione finora
- Anabolismo de Los CarbohidratosDocumento22 pagineAnabolismo de Los CarbohidratosStephanie VallejosNessuna valutazione finora
- En ZimasDocumento2 pagineEn ZimasAdriana RicoNessuna valutazione finora
- TEMA20Documento46 pagineTEMA20Mariana Linares QuevedoNessuna valutazione finora
- Repaso Parciales Bioquímica II-1Documento12 pagineRepaso Parciales Bioquímica II-1Ramón MartínezNessuna valutazione finora
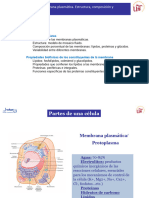
- Tema 2.1. Membrana Plasmática. Estructura, Composición y Propiedades.Documento18 pagineTema 2.1. Membrana Plasmática. Estructura, Composición y Propiedades.pruizi14Nessuna valutazione finora
- 6-7-8. Plaquetas y HemostasiaDocumento22 pagine6-7-8. Plaquetas y HemostasiaNadir GasparNessuna valutazione finora
- TCQ HuevoDocumento2 pagineTCQ HuevoIvanaNessuna valutazione finora
- Tema 15. Biosíntesis de Ácidos GrasosDocumento22 pagineTema 15. Biosíntesis de Ácidos Grasosnestors89Nessuna valutazione finora
- Programa de Bioquímica GeneralDocumento3 paginePrograma de Bioquímica GeneralAngy Carolina Castaño OspinaNessuna valutazione finora
- DislipidemiasDocumento4 pagineDislipidemiasDaniela HerreraNessuna valutazione finora
- Metabolismo de Los LípidosDocumento26 pagineMetabolismo de Los LípidosDiara MaylénNessuna valutazione finora
- Quimica Bioinorganica Del Hierro 1Documento66 pagineQuimica Bioinorganica Del Hierro 1Edward Ernesto Asencios BuriackNessuna valutazione finora
- S7 - L1. Biosíntesis de Ácidos Grasos y Eicosanoides PDFDocumento16 pagineS7 - L1. Biosíntesis de Ácidos Grasos y Eicosanoides PDFGaby Lisbeth LlhNessuna valutazione finora
- Copia de Bioquimica Preguntas Sin ResponderDocumento3 pagineCopia de Bioquimica Preguntas Sin ResponderNoeCQNessuna valutazione finora
- Sintesis Del Palmitato-JosephDocumento7 pagineSintesis Del Palmitato-JosephJoseph Orozco Martinez100% (1)
- Biosíntesis de Ácidos GrasosDocumento3 pagineBiosíntesis de Ácidos GrasosHelen AjuNessuna valutazione finora
- 2do PARCIAL BIOQUIDocumento19 pagine2do PARCIAL BIOQUIgianellaNessuna valutazione finora
- HIDROLASADocumento6 pagineHIDROLASAGUILMER ALMIRON CCORIHUAMANNessuna valutazione finora