Potrebbero piacerti anche
- GUIÓN TÉCNICO PARA VIDEO (Ejemplo)Documento1 paginaGUIÓN TÉCNICO PARA VIDEO (Ejemplo)Ruben Dguez78% (32)
- Sndrome de Respuesta Inflamatoria SistmicaDocumento4 pagineSndrome de Respuesta Inflamatoria SistmicaRosa Mezta DamisNessuna valutazione finora
- Monografia InflamacionDocumento27 pagineMonografia InflamacionJudithGonzálesInga100% (2)
- El Libro de la Dieta Antiinflamatoria: Plan de 14 días para Sanar el Sistema inmunológico y Sentirte Mejor que NuncaDa EverandEl Libro de la Dieta Antiinflamatoria: Plan de 14 días para Sanar el Sistema inmunológico y Sentirte Mejor que NuncaValutazione: 4.5 su 5 stelle4.5/5 (14)
- Decisiones Extremas Ensayo Critico.Documento2 pagineDecisiones Extremas Ensayo Critico.Lala PuertaNessuna valutazione finora
- Respuesta InflamatoriaDocumento4 pagineRespuesta InflamatoriaMariana Bueno GarciaNessuna valutazione finora
- Respuesta Endocrina y Metabólica Al TraumaDocumento7 pagineRespuesta Endocrina y Metabólica Al TraumaJorge GonzalezNessuna valutazione finora
- PATOLOGIA DeberDocumento9 paginePATOLOGIA DeberMircka CedeñoNessuna valutazione finora
- SirsDocumento15 pagineSirsYummyNessuna valutazione finora
- Cascada Inflamatoria e InflamacionDocumento6 pagineCascada Inflamatoria e InflamacionAngelo Maycol P EspinozaNessuna valutazione finora
- Proceso InflamatorioDocumento5 pagineProceso InflamatorioRen VerboonenNessuna valutazione finora
- Proceso InflamatorioDocumento5 pagineProceso InflamatorioDavid Morales ZepedaNessuna valutazione finora
- Fisiopatología de Falla Orgánica MúltipleDocumento9 pagineFisiopatología de Falla Orgánica MúltipleManny BeatzNessuna valutazione finora
- InflamaciónDocumento8 pagineInflamaciónJhon HortealesNessuna valutazione finora
- Inflamación ProDocumento19 pagineInflamación ProDaillynNessuna valutazione finora
- SirsDocumento31 pagineSirsRicardo RománNessuna valutazione finora
- Conferencia Síndrome de Respuesta Inflamatoria SistémicaDocumento44 pagineConferencia Síndrome de Respuesta Inflamatoria SistémicaGenesis PerezNessuna valutazione finora
- Sepsis y Shock SepticoDocumento8 pagineSepsis y Shock SepticoJuan Sebastian ValenciaNessuna valutazione finora
- CAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoDocumento91 pagineCAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoHector Adrian SalasNessuna valutazione finora
- CAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoDocumento91 pagineCAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoAlexis AlonsoNessuna valutazione finora
- INFLAMACIONDocumento5 pagineINFLAMACIONCristian Flores TorricoNessuna valutazione finora
- Cuestionario de INFLAMACIÓNDocumento25 pagineCuestionario de INFLAMACIÓNRodrigo Mata GomezNessuna valutazione finora
- Taller 1 Inmunologia 2 CorteDocumento3 pagineTaller 1 Inmunologia 2 CorteJHON ARGOTENessuna valutazione finora
- Teoría 6 SIRS - SHOCK PDFDocumento27 pagineTeoría 6 SIRS - SHOCK PDFKiara Gonzales PalaciosNessuna valutazione finora
- Resumen Sepsis y Choque SepticoDocumento6 pagineResumen Sepsis y Choque SepticoJuan RodriguezNessuna valutazione finora
- CAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoDocumento91 pagineCAPÍTULO 2 - Respuesta Sistémica A La Lesión y Apoyo MetabólicoMontes InésNessuna valutazione finora
- Homeostasis y CirugíaDocumento8 pagineHomeostasis y CirugíaMaría Alejandra LacayoNessuna valutazione finora
- Conferencia 023 - Síndrome de Respuesta Inflamatoria Sistémica.Documento44 pagineConferencia 023 - Síndrome de Respuesta Inflamatoria Sistémica.YALIELNessuna valutazione finora
- La Interacción Inmuno-neuro-Endocrina en Enfermedades Reumáticas Autoinmunes - Un Nuevo Desafio para El Reumatólogo (2011)Documento3 pagineLa Interacción Inmuno-neuro-Endocrina en Enfermedades Reumáticas Autoinmunes - Un Nuevo Desafio para El Reumatólogo (2011)ALICIANessuna valutazione finora
- PatologiaDocumento32 paginePatologiaselena mujicaNessuna valutazione finora
- 001 Sris y SdmoDocumento6 pagine001 Sris y Sdmovitorinno5705100% (2)
- INFLAMACIONDocumento20 pagineINFLAMACIONDanielaNessuna valutazione finora
- SirsDocumento41 pagineSirsJuan Carlos De la Torre100% (1)
- Inflamación y Reparación TerminadoDocumento29 pagineInflamación y Reparación TerminadoAxel Nuñez100% (2)
- Capítulo 6 - MergedDocumento6 pagineCapítulo 6 - MergedMonserrat Sánchez NietoNessuna valutazione finora
- 001 Sris y SdmoDocumento7 pagine001 Sris y Sdmovitorinno5705Nessuna valutazione finora
- Respuesta Al Ayuno, Inanición y EstrésDocumento4 pagineRespuesta Al Ayuno, Inanición y EstrésMiguelLópezNessuna valutazione finora
- Inflamación y Reparación TisularDocumento20 pagineInflamación y Reparación TisularMaría FernandaNessuna valutazione finora
- SepsisDocumento22 pagineSepsisBreenda CGNessuna valutazione finora
- Fases de LaSepsisDocumento10 pagineFases de LaSepsisDnns PrietNessuna valutazione finora
- Sepsis Fisiopatologia y Avances en El Tratamiento PDFDocumento11 pagineSepsis Fisiopatologia y Avances en El Tratamiento PDFMauricio CarreñoNessuna valutazione finora
- Ensayo de FisiopatologíaDocumento5 pagineEnsayo de FisiopatologíaGema Garcia BravoNessuna valutazione finora
- Capitulo 5 InmunologiaDocumento18 pagineCapitulo 5 InmunologiaDaniel Alberto Garcia AguilarNessuna valutazione finora
- INMUNONUTRICIONDocumento10 pagineINMUNONUTRICIONAndrés QuijanoNessuna valutazione finora
- Respuesta Metabólica Al TraumaDocumento4 pagineRespuesta Metabólica Al Traumadroejj100% (2)
- Alergia SDocumento11 pagineAlergia SSofíaNessuna valutazione finora
- Trauma y SepsisDocumento12 pagineTrauma y SepsisNutriólogo Adar LazaroNessuna valutazione finora
- Caracteristícas de La InflamaciónDocumento8 pagineCaracteristícas de La Inflamaciónfrida ornelasNessuna valutazione finora
- Reaccion AnafilacticaDocumento3 pagineReaccion AnafilacticaManfredo Ricardo Periche ChambaNessuna valutazione finora
- 2.material de Lectura - Semana 2 - Pato QXDocumento19 pagine2.material de Lectura - Semana 2 - Pato QXspeedlokoNessuna valutazione finora
- Capítulo 6Documento4 pagineCapítulo 6Monserrat Sánchez NietoNessuna valutazione finora
- Falla Orgánica MúltipleDocumento8 pagineFalla Orgánica MúltipleGiordan SanchezNessuna valutazione finora
- Quemaduras y Su Respuesta InmunologicaDocumento5 pagineQuemaduras y Su Respuesta InmunologicalocurasdeyoNessuna valutazione finora
- Reacciones Farmacológicas CutáneasDocumento9 pagineReacciones Farmacológicas CutáneasJuan Carlos OrtizNessuna valutazione finora
- Desarrollo de Modulo Iv Críticos SepsisDocumento27 pagineDesarrollo de Modulo Iv Críticos SepsisHéctor Juárez GrandaNessuna valutazione finora
- S0210569100796239 S300 EsDocumento10 pagineS0210569100796239 S300 EsjerejerejereNessuna valutazione finora
- Síndrome de Respuesta Inflamatoria SistémicaDocumento6 pagineSíndrome de Respuesta Inflamatoria Sistémicalucero166Nessuna valutazione finora
- Inflamacion y NutricionDocumento14 pagineInflamacion y NutricionAbigail Estefany Suero PerezNessuna valutazione finora
- Proceso InflamatorioDocumento3 pagineProceso InflamatoriosusanNessuna valutazione finora
- Respuesta InflamatoriaDocumento39 pagineRespuesta InflamatoriaIEIASELNessuna valutazione finora
- Almodóvar Por Lucrecia MartelDocumento2 pagineAlmodóvar Por Lucrecia MartelMijail Miranda ZapataNessuna valutazione finora
- Convocatoria 2020 Festival RadicalDocumento2 pagineConvocatoria 2020 Festival RadicalMijail Miranda ZapataNessuna valutazione finora
- Traducción Conferencia Vandana Shiva PDFDocumento6 pagineTraducción Conferencia Vandana Shiva PDFMijail Miranda ZapataNessuna valutazione finora
- Spinoza Disidente Diego Tatin PDFDocumento2 pagineSpinoza Disidente Diego Tatin PDFMijail Miranda ZapataNessuna valutazione finora
- Preguntas Chaqueo SCZDocumento2 paginePreguntas Chaqueo SCZMijail Miranda ZapataNessuna valutazione finora
- Yawar MallkuDocumento1 paginaYawar MallkuMijail Miranda ZapataNessuna valutazione finora
- Chachawarmi A Lo YungueñoDocumento22 pagineChachawarmi A Lo YungueñoMijail Miranda ZapataNessuna valutazione finora
- PRONUNCIAMIENTO ARAMBURO-EID-OÑA (Premio Peter Travesí 2015)Documento1 paginaPRONUNCIAMIENTO ARAMBURO-EID-OÑA (Premio Peter Travesí 2015)Mijail Miranda ZapataNessuna valutazione finora
- Peter Travesí, Ese Que Vuelve Cada SeptimebreDocumento4 paginePeter Travesí, Ese Que Vuelve Cada SeptimebreMijail Miranda ZapataNessuna valutazione finora
- Reseña Yawar MallkuDocumento2 pagineReseña Yawar MallkuMijail Miranda ZapataNessuna valutazione finora
- Reseña - La Santa Cruz de SadeDocumento3 pagineReseña - La Santa Cruz de SadeMijail Miranda ZapataNessuna valutazione finora
- Edema Pulmonar AgudoDocumento48 pagineEdema Pulmonar AgudoMijail Miranda ZapataNessuna valutazione finora
- Tragedia en Tres Actos-Alfonso SaasteDocumento3 pagineTragedia en Tres Actos-Alfonso SaasteMijail Miranda ZapataNessuna valutazione finora
- Reglamentos de Desarrollo Humano y CulturaDocumento351 pagineReglamentos de Desarrollo Humano y CulturaMijail Miranda ZapataNessuna valutazione finora
- Especialidad Medicina Critica y Terapia IntensivaDocumento2 pagineEspecialidad Medicina Critica y Terapia IntensivavictorhugopedrazaNessuna valutazione finora
- Escala de Estres Ocupacional OitDocumento2 pagineEscala de Estres Ocupacional OitMijael Ptm Aquino BelizarioNessuna valutazione finora
- Siempre AliceDocumento2 pagineSiempre Alicejuanda18Nessuna valutazione finora
- Prolapso RectalDocumento3 pagineProlapso RectalFernando RivasNessuna valutazione finora
- Bordetella Pertussis LetiDocumento9 pagineBordetella Pertussis LetiLeticia Nevado GarciaNessuna valutazione finora
- Guía Miceliales Oportunistas 2016 DIFUSIÓNDocumento138 pagineGuía Miceliales Oportunistas 2016 DIFUSIÓNJavier J Torres PNessuna valutazione finora
- Glosario Sistema Genitourinario 2.0Documento7 pagineGlosario Sistema Genitourinario 2.0eduortizNessuna valutazione finora
- Lidocainacon EpinefrinaDocumento7 pagineLidocainacon EpinefrinamonzukaNessuna valutazione finora
- 5 Tecnicas de Neuroimagen PDFDocumento36 pagine5 Tecnicas de Neuroimagen PDFJohanna Denisse Martínez CaroNessuna valutazione finora
- HIDROCELEDocumento2 pagineHIDROCELERoxana Pozo100% (1)
- 18 Beneficios Del Jengibre para Tu Cuerpo y Tu SaludDocumento11 pagine18 Beneficios Del Jengibre para Tu Cuerpo y Tu SaludMITCHAEL CASASNessuna valutazione finora
- Disfonia InfantilDocumento25 pagineDisfonia InfantilThamara Castillo CisternasNessuna valutazione finora
- Grupos FarmacologicosDocumento95 pagineGrupos FarmacologicosCamilo Bastidas100% (2)
- AVENTHO Tarjetas NuevasDocumento17 pagineAVENTHO Tarjetas NuevasReynaldo Torres Carrillo100% (1)
- 2 Obtención de Muestras SanguíneasDocumento3 pagine2 Obtención de Muestras SanguíneasDaniel Darwin Choque UscaNessuna valutazione finora
- TaquicardiaDocumento12 pagineTaquicardiatomas romeroNessuna valutazione finora
- FarmacometríaDocumento94 pagineFarmacometríaLuisNessuna valutazione finora
- Interdental CAL en Sitio de Mayor Ubicación Perdida de Hueso Radiografica Dientes Perdidos 1 A 2 MM 3 A 4 MM 5 MM 5 MMDocumento2 pagineInterdental CAL en Sitio de Mayor Ubicación Perdida de Hueso Radiografica Dientes Perdidos 1 A 2 MM 3 A 4 MM 5 MM 5 MMJd CruzNessuna valutazione finora
- Certificado de Salud (Leo) Zum Izt 2022Documento1 paginaCertificado de Salud (Leo) Zum Izt 2022Luis Leonardo Rodrigo Franco SánchezNessuna valutazione finora
- Hernia UmbilicalDocumento20 pagineHernia UmbilicalArmandoGomezNessuna valutazione finora
- Guatemala Catalogo de Postgrados USACDocumento16 pagineGuatemala Catalogo de Postgrados USACEve Cano de Lorenzo100% (1)
- Test EnfermeríaDocumento9 pagineTest EnfermeríaSergio Garcia Isabel BerenguelNessuna valutazione finora
- 15.051 Indicaciones de La Biopsia Pleural, Pleuroscopia y MediastinosDocumento3 pagine15.051 Indicaciones de La Biopsia Pleural, Pleuroscopia y MediastinosJose YoveraNessuna valutazione finora
- Ictericia e Insuficiencia RenalDocumento7 pagineIctericia e Insuficiencia RenalYennifer ManriqueNessuna valutazione finora
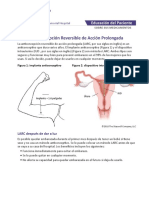
- Northwestern Medicine Anticoncepcion Reversible de Accion Prolongada LarcDocumento2 pagineNorthwestern Medicine Anticoncepcion Reversible de Accion Prolongada LarcOscar Mauricio Ortega CarvajalNessuna valutazione finora
- 15 Sedación y AnalgesiaDocumento37 pagine15 Sedación y AnalgesiamaguisssaNessuna valutazione finora
- Atención Farmacéutica Primera Clase Unica Abril2015Documento32 pagineAtención Farmacéutica Primera Clase Unica Abril2015Nadia PDNessuna valutazione finora
- Sindrome Nefrotico y Nefritico PediatriaDocumento56 pagineSindrome Nefrotico y Nefritico PediatriaHair Alexander Rojas RojasNessuna valutazione finora
- 2019 Mayo 06 ROMERO MERO JOSUE GABRIEL Cert. Notas Global ANUAL 2011-2018IR-Signed-signed-signedDocumento3 pagine2019 Mayo 06 ROMERO MERO JOSUE GABRIEL Cert. Notas Global ANUAL 2011-2018IR-Signed-signed-signedHugo HaroNessuna valutazione finora