Potrebbero piacerti anche
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Respiratory System: " " and Complete The Following ActivitiesDocumento3 pagineThe Respiratory System: " " and Complete The Following ActivitieswenukaNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Jural Keauratan KodeDocumento8 pagineJural Keauratan KodeShela MaranataNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Analisis JurnalDocumento4 pagineAnalisis JurnalSuminar NazirNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Lo MastitisDocumento12 pagineLo MastitisRaul MooreNessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
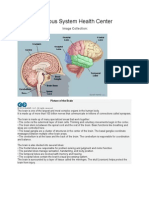
- Brain & Nervous System Health Center: Image Collection: Human AnatomyDocumento1 paginaBrain & Nervous System Health Center: Image Collection: Human AnatomyNica NebrejaNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Thyroid CrisisDocumento2 pagineThyroid CrisisJarelle Guinto RomanNessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Human ReproductionDocumento35 pagineHuman ReproductionMERYL SALOMINA FRANCISCONessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hans - Grade 4 1st PeriodicalsDocumento87 pagineHans - Grade 4 1st PeriodicalsphenorenNessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Tetralogy of FallotDocumento38 pagineTetralogy of FallotJohn Paul MedalloNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Interpretation of FibroScanDocumento16 pagineInterpretation of FibroScanAAMIRNessuna valutazione finora
- 2010 Spasticity MechanismsDocumento10 pagine2010 Spasticity Mechanismsachsyaf arfanNessuna valutazione finora
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- DashBoard Health SampleDocumento1 paginaDashBoard Health SampleAliChana1Nessuna valutazione finora
- De Belen, Anjanette Z - MODULE 7 The SKINDocumento2 pagineDe Belen, Anjanette Z - MODULE 7 The SKINDe Belen, Anjanette ZNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- UEU Dietetik Penyakit Tidak Menular Pertemuan 9Documento32 pagineUEU Dietetik Penyakit Tidak Menular Pertemuan 9Riez Faisal NNessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- INTEGUMENTARY-SYS With AnimationDocumento43 pagineINTEGUMENTARY-SYS With AnimationsattNessuna valutazione finora
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- What Is Nervous System?: Presented By: TIP Grade 12 StudentDocumento28 pagineWhat Is Nervous System?: Presented By: TIP Grade 12 StudentDreiza Patria SunodanNessuna valutazione finora
- Responses To Altered Oxygenation, Cardiac and Tissue - Anatomy To AssessmentDocumento254 pagineResponses To Altered Oxygenation, Cardiac and Tissue - Anatomy To Assessmentlouradel100% (1)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Memorise in A Creative WaysDocumento22 pagineMemorise in A Creative WaysWinnie MasturaNessuna valutazione finora
- Sabiston Textbook of Surgery, 18th Ed. The Biological Basis of Modern Surgical PracticeDocumento5 pagineSabiston Textbook of Surgery, 18th Ed. The Biological Basis of Modern Surgical PracticeNunsio NallaNessuna valutazione finora
- Immune SystemDocumento34 pagineImmune SystemB.simhachalam100% (1)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Male Reproductive Teacher NotesDocumento3 pagineMale Reproductive Teacher Notespaigebotlon30Nessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Midterm Worksheet ALHS 1090 AssignmentDocumento13 pagineMidterm Worksheet ALHS 1090 AssignmentElijah HallNessuna valutazione finora
- Crush InjuryDocumento14 pagineCrush InjuryDioNessuna valutazione finora
- Psychology Unit 3 Aos1 Outcome 1 Aa Aa SMDocumento68 paginePsychology Unit 3 Aos1 Outcome 1 Aa Aa SMapi-298026161Nessuna valutazione finora
- The Child With Endocrine DysfunctionDocumento5 pagineThe Child With Endocrine Dysfunctionhenny1620Nessuna valutazione finora
- Salivary GlandsDocumento97 pagineSalivary GlandsSwati George100% (2)
- Anatomy and Physiology Colouring and Workbook 5th EditionDocumento317 pagineAnatomy and Physiology Colouring and Workbook 5th EditionSergio Alejandro Perez Solis56% (9)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Week 2Documento8 pagineWeek 2Kris Adrian MallillinNessuna valutazione finora
- Physiology of VomittingDocumento23 paginePhysiology of VomittingPrakash PanthiNessuna valutazione finora
- Somatic Nervous SystemDocumento20 pagineSomatic Nervous SystemRichmond Czyrus LopezNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)