Potrebbero piacerti anche
- Modelos Cinetica EnzimaticaDocumento4 pagineModelos Cinetica EnzimaticaDui Dui Dui DuiNessuna valutazione finora
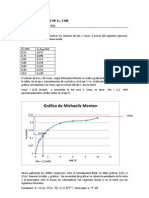
- Ecuación Michaelis-Menten: Modelo cinético enzimáticoDocumento3 pagineEcuación Michaelis-Menten: Modelo cinético enzimáticoLourdes Sánchez100% (1)
- Apuntesparte5 10178Documento38 pagineApuntesparte5 10178Josimar Pasquel100% (1)
- Modelo cinético Michaelis-MentenDocumento3 pagineModelo cinético Michaelis-MentenFiorella Rosas Ruiz100% (1)
- Modelo cinético Michaelis-MentenDocumento7 pagineModelo cinético Michaelis-MentenEdward Jorge Matias AtenciaNessuna valutazione finora
- Bioqimica 5Documento3 pagineBioqimica 5Jose Luis Diaz AyraNessuna valutazione finora
- Cinética EnzimáticaDocumento11 pagineCinética EnzimáticaVirginia Elena Jimenez JimenezNessuna valutazione finora
- Tema 012Documento27 pagineTema 012Luis Salinas ANessuna valutazione finora
- Determinación de los parámetros Vmax y Km de la ecuación de Michaelis-MentenDocumento5 pagineDeterminación de los parámetros Vmax y Km de la ecuación de Michaelis-MentenAndrés Felipe Aristizabal MonterrozaNessuna valutazione finora
- Tema 012Documento27 pagineTema 012Franz Tucta HuillcaNessuna valutazione finora
- Determinaciòn de Los Parámetros Cinéticos de La EnzimaDocumento30 pagineDeterminaciòn de Los Parámetros Cinéticos de La EnzimaLeon VásquezNessuna valutazione finora
- Clase 10Documento37 pagineClase 10Kade CadeNessuna valutazione finora
- Forma Integrada de La Ecuación de MichaelisDocumento5 pagineForma Integrada de La Ecuación de MichaelisAngela CaceresNessuna valutazione finora
- Conceptos Básicos de Cinética QuímicaDocumento8 pagineConceptos Básicos de Cinética QuímicaNicole Del Solar LagosNessuna valutazione finora
- Cinética de Las Reacciones para Sistemas BiológicosDocumento41 pagineCinética de Las Reacciones para Sistemas BiológicosJessy HernándezNessuna valutazione finora
- Determinación de la cinética enzimática de la invertasa mediante el análisis de Michaelis-MentenDocumento6 pagineDeterminación de la cinética enzimática de la invertasa mediante el análisis de Michaelis-MentenDago OrtizNessuna valutazione finora
- CINÉTICA ENZIMÁTICA - Biologia y QuimicaDocumento12 pagineCINÉTICA ENZIMÁTICA - Biologia y QuimicaWILDER PIZARRO CUNYASNessuna valutazione finora
- Laboratorio #2Documento7 pagineLaboratorio #2Carlos César Távara LimasNessuna valutazione finora
- Presentacion 6 Cinetica EnzimaticaDocumento46 paginePresentacion 6 Cinetica EnzimaticaE Clinton LMNessuna valutazione finora
- Diapositivas de Métodos Matemáticos para Calcular Parámetros CinéticosDocumento30 pagineDiapositivas de Métodos Matemáticos para Calcular Parámetros CinéticosMirella Corina Vargas GòmezNessuna valutazione finora
- Actividad Enzimática de La Fosfatasa AlcalinaDocumento16 pagineActividad Enzimática de La Fosfatasa AlcalinaDayana Florez GamboaNessuna valutazione finora
- ProcesosExtraccion LiquidoLiquidoDocumento18 pagineProcesosExtraccion LiquidoLiquidoMiguel RomerosNessuna valutazione finora
- Ingenieria de Reacciones Enzimàticas: Desarrollo de Expresiones Cinèticas.Documento21 pagineIngenieria de Reacciones Enzimàticas: Desarrollo de Expresiones Cinèticas.Balam ToscanoNessuna valutazione finora
- Considere La Reacción de Transformación A Gas de AguaDocumento17 pagineConsidere La Reacción de Transformación A Gas de AguaManuel BloodHoof Merlo100% (2)
- Tutorial Calculos de KM y VmaxDocumento3 pagineTutorial Calculos de KM y VmaxFrancisca Fuentes100% (1)
- Catalisis 2Documento36 pagineCatalisis 2ROSARIOMARTINEZ25Nessuna valutazione finora
- Biocatalisis o Catalisis EnzimaticaDocumento21 pagineBiocatalisis o Catalisis EnzimaticaLuis KrNessuna valutazione finora
- BiofísicaDocumento33 pagineBiofísicaKlaus KöhlerNessuna valutazione finora
- CSTR No Isotermico VictorDocumento6 pagineCSTR No Isotermico Victorzwerner100% (1)
- Cinética de Michalis-Menten y de Briggs-Haldane para La Glucosa IsomerasaDocumento4 pagineCinética de Michalis-Menten y de Briggs-Haldane para La Glucosa IsomerasaMokr Guerrero EsperanzaNessuna valutazione finora
- Tarea 2 Cinética EnzimáticaDocumento4 pagineTarea 2 Cinética EnzimáticaRashell AdamesNessuna valutazione finora
- Ecuaciones de Michaelis PDFDocumento2 pagineEcuaciones de Michaelis PDFAmaury Daniel CortesNessuna valutazione finora
- EnzInhibDocumento14 pagineEnzInhibDavid AntonioNessuna valutazione finora
- ENZIMAS - Biotecnología 2.1Documento78 pagineENZIMAS - Biotecnología 2.1BarbyModestiNessuna valutazione finora
- Deducción Ecuación Michaelis MentenDocumento11 pagineDeducción Ecuación Michaelis Mententere ramirezNessuna valutazione finora
- CEII Tarea2 Equipo6Documento14 pagineCEII Tarea2 Equipo6Rosalino Leonides Martinez MarquezNessuna valutazione finora
- Cinetica de Michaelis' Menten UltimoDocumento5 pagineCinetica de Michaelis' Menten UltimoWilliam Adriano ValleNessuna valutazione finora
- Cuestionario enzimas: energía de activación, equilibrio químico y cinética enzimáticaDocumento5 pagineCuestionario enzimas: energía de activación, equilibrio químico y cinética enzimáticaNestor RuizNessuna valutazione finora
- Biocinética y BiorreactoresDocumento8 pagineBiocinética y Biorreactorestaniamariaguzman_353Nessuna valutazione finora
- Capitulo 8 Metodo de TakabeyaDocumento27 pagineCapitulo 8 Metodo de TakabeyaFrancisco Lucero80% (5)
- Bio Qui MicaDocumento4 pagineBio Qui MicaAlejandra SalcedoNessuna valutazione finora
- Problemas de EnzimologíaDocumento24 pagineProblemas de EnzimologíasofiaNessuna valutazione finora
- Aplicaciones Del Ensayo TriaxialDocumento31 pagineAplicaciones Del Ensayo Triaxialrobertok1704Nessuna valutazione finora
- Informe Tirosinasa 2 y 3 PDFDocumento6 pagineInforme Tirosinasa 2 y 3 PDFAndre CadavidNessuna valutazione finora
- Soluciones: Capítulo 5 Propiedades de Las EnzimasDocumento5 pagineSoluciones: Capítulo 5 Propiedades de Las EnzimasAngelesNessuna valutazione finora
- Modelo Matemático Del Flujo IsotérmicoDocumento7 pagineModelo Matemático Del Flujo IsotérmicoMcGruber_CNessuna valutazione finora
- CAPITULO12Documento38 pagineCAPITULO12Jenny Liberato FloresNessuna valutazione finora
- Ejercicio Cinetica Fabian Franco VDocumento6 pagineEjercicio Cinetica Fabian Franco VfabianfravalNessuna valutazione finora
- Metodo Newton RaphsonDocumento4 pagineMetodo Newton RaphsonAndrea NuñezNessuna valutazione finora
- Cinetica EnzimáticaDocumento38 pagineCinetica EnzimáticaFernanda Castillo ÜNessuna valutazione finora
- Taller de Bioquimica EnzimasDocumento15 pagineTaller de Bioquimica EnzimasAlejandra SalcedoNessuna valutazione finora
- 4.2.2 Modelo de Michaellis MenteDocumento11 pagine4.2.2 Modelo de Michaellis MenteDiana Akatzukis Gusemëz50% (2)
- Cinética Enzimática PDFDocumento13 pagineCinética Enzimática PDFDurvelito Chulo GuapoNessuna valutazione finora
- Taller 2 Producción de SO3Documento5 pagineTaller 2 Producción de SO3jtabaresh100% (1)
- Ejercicios de Integrales y Ecuaciones Integrales-DiferencialesDa EverandEjercicios de Integrales y Ecuaciones Integrales-DiferencialesNessuna valutazione finora
- Conan 16Documento139 pagineConan 16Mario LanillaNessuna valutazione finora
- Separata ApiculturaDocumento60 pagineSeparata ApiculturaAlonso Lopez MendozaNessuna valutazione finora
- Referencias Normativas para Procesos LácteosDocumento5 pagineReferencias Normativas para Procesos Lácteosingeniero035Nessuna valutazione finora
- NadaDocumento4 pagineNadaAnonymous J7wT7i7Nessuna valutazione finora
- Neumonia Afebril Del LactanteDocumento4 pagineNeumonia Afebril Del LactanteLAURA CATALINA AGUIRRE SANTOS100% (1)
- Músculo CardiacoDocumento50 pagineMúsculo CardiacoMilagros Mariños RodriguezNessuna valutazione finora
- Síndrome de Apnea-Hipopnea Del SueñoDocumento11 pagineSíndrome de Apnea-Hipopnea Del SueñoJimmy Sulca CorreaNessuna valutazione finora
- Comparativa organelos celularesDocumento3 pagineComparativa organelos celularesRosy Vargas Caro0% (2)
- Criptorquidia GPC SsaDocumento35 pagineCriptorquidia GPC SsaLuis SanzNessuna valutazione finora
- PENTOBARBITALDocumento4 paginePENTOBARBITALPaola Salas CáceresNessuna valutazione finora
- Arancel Fonasa Actualizado 31 08 2011Documento31 pagineArancel Fonasa Actualizado 31 08 2011Cristian ValdesNessuna valutazione finora
- Síndromes en Medicina Tradicional China, Que Se Asocian Con Dolor de Hombro. Propuestas de Manejo Con Acupuntura y MoxibustiónDocumento30 pagineSíndromes en Medicina Tradicional China, Que Se Asocian Con Dolor de Hombro. Propuestas de Manejo Con Acupuntura y MoxibustiónJuanNessuna valutazione finora
- 2-QUÍMICA (7 - 12) Corr Nuevo OkDocumento49 pagine2-QUÍMICA (7 - 12) Corr Nuevo Oklenin churaNessuna valutazione finora
- AuriculoterapiaDocumento46 pagineAuriculoterapiaSaffiah Calcagno100% (1)
- SISTEMA CIRCULATORIO AREA C y TDocumento6 pagineSISTEMA CIRCULATORIO AREA C y TKarina M. NoaNessuna valutazione finora
- Caballero - Manual para Educar La Voz Hablada y CantadaDocumento21 pagineCaballero - Manual para Educar La Voz Hablada y CantadaceregalNessuna valutazione finora
- Fibromialgia y Fisioterapia Global PDFDocumento5 pagineFibromialgia y Fisioterapia Global PDFTephi Gomez ImaraiNessuna valutazione finora
- Terminología Educación para La SaludDocumento8 pagineTerminología Educación para La SaludNatalia Sánchez100% (2)
- Informe Corpúsculo de BarrDocumento9 pagineInforme Corpúsculo de BarrJorge Blanquiceth100% (1)
- 2 Series para El Campo ElectromagnéticoDocumento4 pagine2 Series para El Campo ElectromagnéticoFUMETA100% (1)
- ElectroquimicaDocumento3 pagineElectroquimicasofiaNessuna valutazione finora
- Sepsis en PediatríaDocumento20 pagineSepsis en PediatríaKarenn GallardooNessuna valutazione finora
- La Fotosintesis TripticoDocumento3 pagineLa Fotosintesis TripticoHoshino Lizzy Kuroineko Mikado63% (8)
- Desarrollo Motor Normal 0-6 MesesDocumento46 pagineDesarrollo Motor Normal 0-6 MeseschinocauNessuna valutazione finora
- MSP Trastornos Hipertensivos Del Embarazo Con Portada 3Documento81 pagineMSP Trastornos Hipertensivos Del Embarazo Con Portada 3Yandry Holguin SabandoNessuna valutazione finora
- Bacterias 2008 Octopus MayaDocumento1 paginaBacterias 2008 Octopus MayaHonorioNessuna valutazione finora
- Leucemias AgudasDocumento4 pagineLeucemias AgudasMigue ChavezNessuna valutazione finora
- Trabjo de Angela de BiologiaDocumento7 pagineTrabjo de Angela de BiologiaMarcos GutierrezNessuna valutazione finora
- Parto DistócicoDocumento31 pagineParto DistócicoFred Ry LCNessuna valutazione finora
- Tablas Psicobio. DrogadiccionDocumento26 pagineTablas Psicobio. DrogadiccionAnka NistreaNessuna valutazione finora