Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
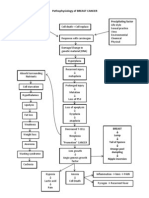
- Pathophysiology of BREAST CANCERDocumento1 paginaPathophysiology of BREAST CANCERAlinor Abubacar100% (6)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Richard D. Lane - Cognitive Neuroscience of EmotionDocumento446 pagineRichard D. Lane - Cognitive Neuroscience of Emotionnexus1968100% (6)
- Medico Legal Aspects of DeathDocumento206 pagineMedico Legal Aspects of Deathrosana f.rodriguez83% (12)
- Biological Level of Analysis Research GuideDocumento45 pagineBiological Level of Analysis Research GuidePhiline Everts100% (2)
- Essentials Lab Resource Guide 12 22Documento67 pagineEssentials Lab Resource Guide 12 22Aijeleth Shahar Gunay AwacayNessuna valutazione finora
- Chapman System of ClassificationDocumento6 pagineChapman System of Classificationvineetvishal73Nessuna valutazione finora
- NURS1603 Course Outline PDFDocumento19 pagineNURS1603 Course Outline PDFYip Ka YiNessuna valutazione finora
- 08 DN 005 BA Int OkDocumento64 pagine08 DN 005 BA Int OkvbogachevNessuna valutazione finora
- DepresijaDocumento6 pagineDepresijaChristopher PhillipsNessuna valutazione finora
- Enzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationDocumento35 pagineEnzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationAkash SinghNessuna valutazione finora
- School: San Agustin Elementary School Grade Level: Six Teacher: Jonalvin A. Ke Learning Area: Science Date: SEPTEMBER 5, 2018 (WEDNESDAY)Documento3 pagineSchool: San Agustin Elementary School Grade Level: Six Teacher: Jonalvin A. Ke Learning Area: Science Date: SEPTEMBER 5, 2018 (WEDNESDAY)Jonalvin KENessuna valutazione finora
- Jump Height Loss As An Indicator of Fatigue During Sprint TrainingDocumento11 pagineJump Height Loss As An Indicator of Fatigue During Sprint TrainingLevyNessuna valutazione finora
- Child Development A Thematic Approach 6th Edition Bukatko Test BankDocumento36 pagineChild Development A Thematic Approach 6th Edition Bukatko Test Bankunframecizarsidquu100% (21)
- Skeletal Muscle Classification and StructureDocumento43 pagineSkeletal Muscle Classification and StructureHusnain WattoNessuna valutazione finora
- MEDICAL SURGICAL NURSING REVIEWDocumento7 pagineMEDICAL SURGICAL NURSING REVIEWeloisa mae gementizaNessuna valutazione finora
- Burn OutDocumento8 pagineBurn OutAtiqahAzizanNessuna valutazione finora
- Fermentation Process KineticsDocumento7 pagineFermentation Process KineticsDillip_subuNessuna valutazione finora
- 7/23/2016 Cristina S. Nebres Mindanao State University at Naawan 1Documento32 pagine7/23/2016 Cristina S. Nebres Mindanao State University at Naawan 1Laila UbandoNessuna valutazione finora
- Trimestral Exam First GradeDocumento3 pagineTrimestral Exam First Gradeemmanuel espinozaNessuna valutazione finora
- Respiratory Muscle Ultrasonography: Methodology, Basic and Advanced Principles and Clinical Applications in ICU and ED Patients-A Narrative ReviewDocumento12 pagineRespiratory Muscle Ultrasonography: Methodology, Basic and Advanced Principles and Clinical Applications in ICU and ED Patients-A Narrative Reviewalejandro RodriguezNessuna valutazione finora
- B.sc. Biotechnology - CcssDocumento61 pagineB.sc. Biotechnology - CcsssreyasbkNessuna valutazione finora
- 5 Day Bodybuilding Workout ScheduleDocumento4 pagine5 Day Bodybuilding Workout ScheduleNazri Yusoff100% (1)
- New European Guidelines Address Hyponatremia ManagementDocumento5 pagineNew European Guidelines Address Hyponatremia ManagementGherciuChirilaLarisaNessuna valutazione finora
- ADocumento2 pagineAイ ロNessuna valutazione finora
- Diagram Human Heart Kel 1Documento3 pagineDiagram Human Heart Kel 1Rahma SafitriNessuna valutazione finora
- Blood ? Summary PDFDocumento10 pagineBlood ? Summary PDFmanar aleneziNessuna valutazione finora
- 0010 (1) Science - Terms 1.154KB PDFDocumento236 pagine0010 (1) Science - Terms 1.154KB PDFMuhammad SaleemNessuna valutazione finora
- Fundamental PositionsDocumento35 pagineFundamental PositionsVinod Kumar50% (4)
- Antiarrhythmic Drugs-Test-1-QuestionsDocumento7 pagineAntiarrhythmic Drugs-Test-1-QuestionsDrishya Bioplannet100% (2)
- CJR Biology UmumDocumento15 pagineCJR Biology UmumGresia FalentinaNessuna valutazione finora