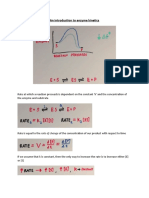
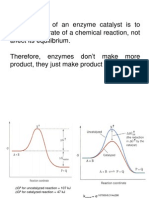
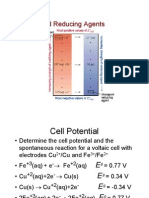
Potrebbero piacerti anche
- Biochem ConceptsDocumento9 pagineBiochem Conceptsdanena88Nessuna valutazione finora
- Enzyme NotesDocumento18 pagineEnzyme NotesDBPNessuna valutazione finora
- Enzyme Catalysis PDFDocumento17 pagineEnzyme Catalysis PDFuzzal ahmedNessuna valutazione finora
- Enzymes - 26th Dec 2022Documento47 pagineEnzymes - 26th Dec 2022Tumpy KumarNessuna valutazione finora
- Part 4 Case Study - Industrial EnzymesDocumento33 paginePart 4 Case Study - Industrial EnzymesyahmedpersNessuna valutazione finora
- Enzyme KineticsDocumento10 pagineEnzyme KineticsQuenneBelocuraNessuna valutazione finora
- Enzyme Kinetics: Medical Biochemistry, Lecture 24Documento39 pagineEnzyme Kinetics: Medical Biochemistry, Lecture 24jeyankarunanithiNessuna valutazione finora
- Chapter 6 NotesDocumento12 pagineChapter 6 NotesCarleyNessuna valutazione finora
- Dokumen - Tips - Enzymes Problem SetDocumento22 pagineDokumen - Tips - Enzymes Problem SetDawlat SlamaNessuna valutazione finora
- Chapter 6 - The Behavior of Proteins ENZYMES-2018Documento111 pagineChapter 6 - The Behavior of Proteins ENZYMES-2018ruaa mhmadNessuna valutazione finora
- Enzyme Kinetics Mechanism and InhibitionDocumento38 pagineEnzyme Kinetics Mechanism and InhibitionManoj SigdelNessuna valutazione finora
- Enzyme Kinetics Practical For RecordingDocumento22 pagineEnzyme Kinetics Practical For RecordingSanya chauhanNessuna valutazione finora
- V VI EnzymesDocumento59 pagineV VI Enzymesthamizh555100% (2)
- Enzymes Kinetics & RegulationDocumento41 pagineEnzymes Kinetics & RegulationConorTankGorbatsjovNessuna valutazione finora
- Science of Living System (BS20001) : - Soumya deDocumento50 pagineScience of Living System (BS20001) : - Soumya deMayank PriayadarshiNessuna valutazione finora
- ERT211 CHP 1-4 PDFDocumento24 pagineERT211 CHP 1-4 PDFsobiyamaragathavelNessuna valutazione finora
- Enz KineticsDocumento20 pagineEnz KineticsSirsha PattanayakNessuna valutazione finora
- Enzymes All NUBDocumento9 pagineEnzymes All NUBPIH SHTNessuna valutazione finora
- 2 Factors Aff & KM, MMeq, DRP 2021Documento29 pagine2 Factors Aff & KM, MMeq, DRP 2021Srishti GoenkaNessuna valutazione finora
- Biology Biomolecules Part 2Documento14 pagineBiology Biomolecules Part 2Ralph Kenneth LunaNessuna valutazione finora
- Energy, Catalysis and Biosynthesis Lecture NotesDocumento17 pagineEnergy, Catalysis and Biosynthesis Lecture Notesmina catNessuna valutazione finora
- Enzymes All NUBDocumento8 pagineEnzymes All NUBPIH SHTNessuna valutazione finora
- Lecture 2 Enzyme KineticsDocumento31 pagineLecture 2 Enzyme KineticsCarinaJongLeeNessuna valutazione finora
- Enzymes - PPT 1Documento54 pagineEnzymes - PPT 1Cesar Augusto Airampo Macedo100% (1)
- Science of Living System: Arindam MondalDocumento47 pagineScience of Living System: Arindam MondalSohini RoyNessuna valutazione finora
- LineweaverBurk PlotDocumento17 pagineLineweaverBurk PlotJawadNessuna valutazione finora
- Enzyme KineticsDocumento10 pagineEnzyme Kineticskedirwoliy100% (1)
- Tugas 2 BiokatalisisDocumento7 pagineTugas 2 BiokatalisisKirstie ImeldaNessuna valutazione finora
- Catalysis and Enzyme Kinetics PDFDocumento43 pagineCatalysis and Enzyme Kinetics PDFBlessings Chawinga100% (1)
- BS 3Documento60 pagineBS 3himanshu_agraNessuna valutazione finora
- Enzyme KineticsDocumento90 pagineEnzyme KineticsAmrit LalNessuna valutazione finora
- Key Topics:: EnzymesDocumento33 pagineKey Topics:: EnzymesMeth100% (1)
- Lec 7 and 8 (Ch. 5) Enz KinDocumento37 pagineLec 7 and 8 (Ch. 5) Enz KinRamy El-HadadNessuna valutazione finora
- IIPE Class 4 24.03Documento41 pagineIIPE Class 4 24.03bharatNessuna valutazione finora
- Enzyme KineticsDocumento19 pagineEnzyme KineticsParinita Mitchelle MandhyanNessuna valutazione finora
- Enzyme and Acid - Base CatalysisDocumento64 pagineEnzyme and Acid - Base Catalysisbinseung skzNessuna valutazione finora
- Lecture 7-SOLS 2023Documento36 pagineLecture 7-SOLS 2023Arghadeep DasNessuna valutazione finora
- Enzyme Kinetics Lab ManualDocumento6 pagineEnzyme Kinetics Lab ManualRudshut CudleNessuna valutazione finora
- Enzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan UniversityDocumento58 pagineEnzyme-Kinetics and Specificity: Modified by Chung-Ming Lin Department of Biotechnology Ming Chuan University陳家琪Nessuna valutazione finora
- Enzyme Part 1-5 Vikneswaran 260110132004Documento7 pagineEnzyme Part 1-5 Vikneswaran 260110132004Vikneswaran VîçkýNessuna valutazione finora
- Enzyme, M-I 024Documento67 pagineEnzyme, M-I 024daman dhamiNessuna valutazione finora
- Enzyme KineticsDocumento13 pagineEnzyme KineticsKhushbu JainNessuna valutazione finora
- Dr. Nazir Ahmad: Nehal BilalDocumento15 pagineDr. Nazir Ahmad: Nehal BilalAayat MughalNessuna valutazione finora
- Enzyme-Catalyst of LifeDocumento7 pagineEnzyme-Catalyst of LifeIrvan DahlanNessuna valutazione finora
- 2023-08-17 EnzymesDocumento42 pagine2023-08-17 EnzymesAjay MahalkaNessuna valutazione finora
- Enzyme2 2012Documento45 pagineEnzyme2 2012Surya Prakash KabiNessuna valutazione finora
- 01 Enzyme KineticsDocumento5 pagine01 Enzyme KineticsmwegiNessuna valutazione finora
- 10-11, Kinetics of Enzyme Catalyzed ReactionDocumento34 pagine10-11, Kinetics of Enzyme Catalyzed ReactionS. Ansary100% (1)
- CH 7 Kinetics and RegulationDocumento8 pagineCH 7 Kinetics and Regulationcococat015Nessuna valutazione finora
- Lecture 3 Notes - EnzymologyDocumento9 pagineLecture 3 Notes - EnzymologyYi Gong100% (3)
- Biochemistry NotesDocumento14 pagineBiochemistry NotesJocabed RadillaNessuna valutazione finora
- Revised Chap13-LectDocumento58 pagineRevised Chap13-LectSimon ChowNessuna valutazione finora
- Enzimas 2015 01 BqaBasicaDocumento128 pagineEnzimas 2015 01 BqaBasicaEdithaSorNessuna valutazione finora
- Chapter 7.7 Abbreviated Lecture NotesDocumento27 pagineChapter 7.7 Abbreviated Lecture NotesZach MaxwellNessuna valutazione finora
- Enzyme KineticsDocumento23 pagineEnzyme Kinetics1DS19CH011 Jashwanth C RNessuna valutazione finora
- ENK Enzyme Kinetics Lab ManualDocumento11 pagineENK Enzyme Kinetics Lab ManualVanesha AnesNessuna valutazione finora
- Biochem Lecture Week 4Documento9 pagineBiochem Lecture Week 4api-178321548Nessuna valutazione finora
- PhysBiochem Enzyme KineticsDocumento29 paginePhysBiochem Enzyme KineticsMohannad WalyNessuna valutazione finora
- Chemical Kinetics & Equilibrium: Essential Chemistry Self-Teaching GuideDa EverandChemical Kinetics & Equilibrium: Essential Chemistry Self-Teaching GuideNessuna valutazione finora
- Jobs MethodDocumento3 pagineJobs MethodKasunBuddika50% (2)
- Complex Ion EquilibriaDocumento6 pagineComplex Ion EquilibriaKasunBuddikaNessuna valutazione finora
- DNA RNA ProteinSynDocumento8 pagineDNA RNA ProteinSynKasunBuddikaNessuna valutazione finora
- Ether Synthesis & ReactionsDocumento3 pagineEther Synthesis & ReactionsKasunBuddikaNessuna valutazione finora
- Binary CodeDocumento3 pagineBinary CodeHaridass DuraiNessuna valutazione finora
- 121 Tutorial How Det Formal Charges SP 2006Documento4 pagine121 Tutorial How Det Formal Charges SP 2006KasunBuddikaNessuna valutazione finora
- Presentation 17Documento38 paginePresentation 17RIFAT ALINessuna valutazione finora
- Introduction To Computing - 1Documento34 pagineIntroduction To Computing - 1KasunBuddikaNessuna valutazione finora
- An Introduction To ComputingDocumento157 pagineAn Introduction To Computingmjanus13100% (1)
- Nernst Equation 3Documento20 pagineNernst Equation 3KasunBuddikaNessuna valutazione finora
- The Nernst EquationDocumento3 pagineThe Nernst Equationsidharth_sNessuna valutazione finora
- Nernst Equation Example Problems: Echem - MCD 1/3/02Documento9 pagineNernst Equation Example Problems: Echem - MCD 1/3/02sparkshockNessuna valutazione finora
- Welcome To Tintagel - A Visitor's GuideDocumento36 pagineWelcome To Tintagel - A Visitor's GuideClerk To The CouncilNessuna valutazione finora
- Different Types of BarsDocumento5 pagineDifferent Types of BarsDekzie Flores MimayNessuna valutazione finora
- Math 22 TQs - FinalsDocumento7 pagineMath 22 TQs - FinalsZaid Hassan GajesNessuna valutazione finora
- Conde Nast 2014 03 UK PDFDocumento180 pagineConde Nast 2014 03 UK PDFAntonio SNessuna valutazione finora
- Shamim & Company: Internship ReportDocumento19 pagineShamim & Company: Internship ReportMUHAMMAD HARIS AMINNessuna valutazione finora
- Promotional Scheme of Amul Ice Cream: A Project Research Report ONDocumento104 paginePromotional Scheme of Amul Ice Cream: A Project Research Report ONAtul MishraNessuna valutazione finora
- Synthetic Membranes For Membrane ProcessesDocumento14 pagineSynthetic Membranes For Membrane ProcessesRishab SharmaNessuna valutazione finora
- Himanshu ProjectDocumento55 pagineHimanshu ProjectrajNessuna valutazione finora
- Madagascar Comoros Islands v1 m56577569830503995Documento15 pagineMadagascar Comoros Islands v1 m56577569830503995Pablo SulicNessuna valutazione finora
- Madam PuddifootsDocumento9 pagineMadam Puddifootserica milaNessuna valutazione finora
- Antonio Pigafetta - First Voyage Around The WorldDocumento8 pagineAntonio Pigafetta - First Voyage Around The WorldKim CatherineNessuna valutazione finora
- The Christmas SongDocumento11 pagineThe Christmas SongRobin H. WilliamsNessuna valutazione finora
- Food & Beverage TrendsDocumento50 pagineFood & Beverage TrendsAntonin LapresleNessuna valutazione finora
- Hort 366 Objective QuestionsDocumento10 pagineHort 366 Objective Questionsmana khantNessuna valutazione finora
- Departure 201-210Documento59 pagineDeparture 201-210Donela Joy AbellanoNessuna valutazione finora
- t036 Past Perfect Past Tense PDFDocumento2 paginet036 Past Perfect Past Tense PDFMarlene DietrixNessuna valutazione finora
- Buku Heritage JamuDocumento105 pagineBuku Heritage JamuHyde BurkeNessuna valutazione finora
- The Celestial AgricultureDocumento10 pagineThe Celestial AgricultureGus TijerinaNessuna valutazione finora
- H Rider Haggard - King Solomon's MinesDocumento138 pagineH Rider Haggard - King Solomon's MinesKAWNessuna valutazione finora
- Research PaperDocumento21 pagineResearch PaperAviaNessuna valutazione finora
- Health Benefits of LycheeDocumento24 pagineHealth Benefits of LycheeMD AbdullahNessuna valutazione finora
- Down Memory LaneDocumento48 pagineDown Memory LanePriyanka GhoshNessuna valutazione finora
- Entrep Business PlanDocumento16 pagineEntrep Business Planmich libreaNessuna valutazione finora
- Alcohol Explained: by William PorterDocumento37 pagineAlcohol Explained: by William PorterDansNessuna valutazione finora
- K To 12 AralinDocumento3 pagineK To 12 Aralinalvin balisbisNessuna valutazione finora
- Study of Encapsulation TechnologyDocumento18 pagineStudy of Encapsulation Technologydevashri_ranadeNessuna valutazione finora
- Doctor Magnethands: A Game For Three or More Drunk PeopleDocumento2 pagineDoctor Magnethands: A Game For Three or More Drunk PeopleTwogunkidNessuna valutazione finora
- Free English Grammar Ebook BeginnerDocumento44 pagineFree English Grammar Ebook BeginnerValdir FrançaNessuna valutazione finora
- Procedure TextDocumento12 pagineProcedure TextBuds PantunNessuna valutazione finora
- Nasikebuli: (Influenced by Arab Culture)Documento3 pagineNasikebuli: (Influenced by Arab Culture)Dicky Surya IINessuna valutazione finora